Cheat varaq: organizmlarning individual rivojlanishi (ontogenez). Ontogenez tushunchasi
Ontogenez(yunoncha cntos - mavjud) yoki individual rivojlanish - zigota yoki boshqa embrion paydo bo'lgan paytdan boshlab uning hayot tsiklining tabiiy tugaguniga qadar (o'limgacha yoki avvalgi qobiliyatida mavjudligini to'xtatgunga qadar) shaxsning rivojlanishi. Genetika nuqtai nazaridan, ontogenez - bu jinsiy hujayralarga kiritilgan irsiy ma'lumotni ochish va amalga oshirish jarayoni.
Ontogenez har qanday shaxsning tizimli mansubligidan qat'iy nazar ajralmas xususiyatdir. Ontogenez paydo bo'lmaganda, hayotning evolyutsiyasini tasavvur qilib bo'lmaydi. Organizmlarning individual rivojlanishi tarixiy rivojlanish bilan chambarchas bog'liq - filogeniya(yunoncha phyle - qabiladan).
Turli xil turlarning individlarining ontogenezi differensiatsiyaning davomiyligi, tezligi va tabiati bilan farqlanadi. Ko'p hujayrali hayvonlar va odamlarda ontogenezning boshlanishidan oldin davr keladi proembrionik (embriondan oldingi) rivojlanish - progenez . Bu davrda jinsiy hujayralar hosil bo'ladi, urug'lanish jarayoni va zigota hosil bo'ladi.
Ontogenezda to'rtta davr mavjud: embriondan oldingi, embrion (prenatal ), postembrionik (tug'ruqdan keyingi ) Va kattalar holati shu jumladan qarish va o'lim. Hayvonlarda embrion davri odatda differensiatsiyaga, o'simliklarda esa postembrional davrga boy bo'ladi. Bu ontogenez davrlarining har biri, o'z navbatida, ketma-ket sifat bosqichlariga bo'linishi mumkin.
Preembrional gametogenez va urug'lanishni o'z ichiga oladi.
Embrion davr tashqi muhitda yoki onaning tanasining reproduktiv tizimida embrionning rivojlanishi va morfogenezning tez jarayonlari bilan tavsiflanadi. Ushbu jarayonlar natijasida qisqa vaqt ichida ko'p hujayrali organizm paydo bo'ladi.
Inson embrion rivojlanishining uchta davri mavjud: boshlang'ich , embrion , homila (homila ).
Boshlang'ich davr embrion rivojlanishining birinchi haftasini qamrab oladi. U urug'lantirilgan paytdan boshlab boshlanadi va embrion bachadon shilliq qavatiga joylashtirilgunga qadar davom etadi.
Embrion odamlarda davr implantatsiya paytidan organogenez jarayoni tugaguniga qadar (2-8 hafta) boshlanadi. Bu davr organogenez jarayonlari, ovqatlanishning o'ziga xos xususiyatlari bilan tavsiflanadi - histiotrofik ovqatlanish, embrion bachadon bezlari sekretsiyasi va bachadon shilliq qavati to'qimalarining parchalanish mahsulotlari bilan oziqlanganda. Rivojlanishning ushbu davrida uzoq vaqt davomida platsenta qon aylanishi mavjud emas va inson embrioniga xos bo'lgan xarakterli xususiyatlar sotib olinadi.
Homila, yoki inson embrion rivojlanishining homila davri, urug'lantirilgandan keyin 9-haftadan boshlanadi va tug'ilishgacha davom etadi. Bu davr o'sishning kuchayishi, jadal rivojlanish jarayonlari va o'ziga xos ovqatlanish xususiyatlari bilan tavsiflanadi - gemotrofik platsenta qon aylanishining ishlashi bilan bog'liq bo'lgan oziqlanish. Inson embrion rivojlanishi davrlarining xususiyatlari 5-jadvalda keltirilgan .
5-jadval
Inson embrioni rivojlanish davrlarining xususiyatlari
Postembrional odamlarda va sutemizuvchilarda davr tug'ilishdan boshlab, embrion membranalardan hayotning oxirigacha boshlanadi va balog'at yoshiga qadar davom etadi. Tuxumdor hayvonlarda bu davr yosh individ tuxum qobig'idan chiqqan paytdan boshlanadi; o'simliklarda - birlamchi ildiz paydo bo'lgan paytdan boshlab.
ga o'tish kattalar tanasi bevosita yoki bilvosita amalga oshirilishi mumkin. Shu munosabat bilan ontogenezning uch turi ajratiladi: lichinka , lichinka bo'lmagan Va intrauterin .
Lichinka, yoki bilvosita Rivojlanishning bu turi ko'plab koelenteratlar, qurtlar, mollyuskalar, qisqichbaqasimonlar, hasharotlar, lanselletlar, o'pka baliqlari va ba'zi suyakli baliqlar va amfibiyalarga xosdir. Rivojlanishning bu turi lichinka bosqichlarining mavjudligi bilan ajralib turadi. Tuxumdan chiqqandan so'ng, lichinkalar faol hayot tarzini olib boradi va o'zlari oziq-ovqat oladi. Lichinkalar ota-ona shakliga o'xshamaydi - ular tuzilishi jihatidan ancha sodda, vaqtinchalik organlarga ega, ular keyinchalik so'riladi (so'riladi) va kattalarda kuzatilmaydi.
Keyingi transformatsiya - metamorfoz - lichinkalarni kattalarga aylantirish turiga qarab amalga oshirilishi mumkin to'liq transformatsiya , bunda lichinka kattalarnikidan keskin farq qiladi va bir qancha rivojlanish bosqichlaridan o'tadi, ularning asosiysi qo'g'irchoq bosqichidir (kapalak). Yoki rivojlanish pupa bosqichisiz sodir bo'ladi - turiga ko'ra to'liq bo'lmagan transformatsiya , va lichinkaning o'zi katta yoshli hayvonga o'xshaydi, lekin hajmi kichikroq (chigirtka, chigirtka).
Lichinka bo'lmagan (Streyt ) rivojlanish turi kattalar ota-ona shakliga o'xshash organizmning paydo bo'lishi bilan tavsiflanadi, lekin undan kichikroq hajmi va to'liq rivojlanmagan reproduktiv apparati bilan farqlanadi. Hayvonlarning bunday shakllarida (baliqlar, sudralib yuruvchilar, qushlar, tuxum qo'yuvchi sut emizuvchilar, bosh oyoqlilar, koelenteratlar) barcha organlar rivojlanishning embrional davrida, o'sish, balog'atga etishish va funktsiyalarning differentsiatsiyasi postembrional davrda sodir bo'ladi. To'g'ridan-to'g'ri rivojlanish tuxumda ko'p miqdorda sarig'i va rivojlanayotgan embrion uchun himoya vositalarining mavjudligi yoki onaning tanasida embrionning rivojlanishi bilan bog'liq.
Intrauterin (Streyt ) filogenetik nuqtai nazardan eng yangi rivojlanish turidir. Bu yuqori sutemizuvchilar va odamlarga xos bo'lib, unda tuxumlar sarig'ida kambag'al bo'lib, embrionning rivojlanishi onaning tanasining bachadonida sodir bo'ladi. Bunday holda, vaqtinchalik ekstraembrion organlar hosil bo'ladi, ularning eng muhimi platsentadir.
Organizmlarning hayot davrlari
Hayot davrasi, yoki rivojlanish tsikli, tananing eng muhim, asosiy holatini belgilovchi ketma-ket bosqichlardan (ko'pincha bosqichlar deb ataladi) iborat - kelib chiqishi , rivojlanish Va ko'payish .
Jinsiy ko'payadigan organizmlarning hayot aylanishlarida ikki bosqich mavjud: gaploid Va diploid . Bu fazalarning nisbiy davomiyligi tirik organizmlarning turli guruhlari vakillari orasida farq qiladi. Shunday qilib, protozoa va zamburug'larda gaploid faza, yuqori o'simliklar va hayvonlarda esa diploid faza ustunlik qiladi.
Evolyutsiya jarayonida diplofazaning uzayishi gaploid holatga nisbatan diploid holatning afzalliklari bilan izohlanadi. Geterozigotalik va retsessivlik tufayli turli xil allellar saqlanib qoladi va diploid holatda to'planadi. Bu populyatsiyalar va turlarning genofondidagi genetik ma'lumotlarning miqdorini oshiradi, bu esa keyingi evolyutsiya uchun istiqbolli irsiy o'zgaruvchanlik zaxirasining shakllanishiga olib keladi. Shu bilan birga, geterozigotalarda zararli retsessiv allellar fenotipning rivojlanishiga ta'sir qilmaydi va organizmlarning hayotiyligini pasaytirmaydi.
Hayotiy davrlar mavjud oddiy Va murakkab . Murakkab bo'lganlar oddiy tsikllardan iborat bo'lib, ular bu holda murakkab tsikldagi ochiq havolalarga aylanadi.
Avlodlarning almashinishi deyarli barcha evolyutsion rivojlangan suv o'tlari va barcha yuqori o'simliklar uchun xarakterlidir. Avlodlar almashinishi kuzatiladigan o'simlikning hayot aylanishining umumlashtirilgan diagrammasi rasmda keltirilgan. o'n bir.
Guruch. 11. O'zaro almashinadigan avlodlarga ega bo'lgan o'simlikning hayot aylanishining umumlashtirilgan sxemasi
Oddiy tsiklli o'simliklarga misol qilib, faqat sporalar bilan ko'payadigan bir hujayrali yashil suv o'tlari Chlorella hisoblanadi. Xlorellalarning rivojlanishi avtosporalardan boshlanadi. Hali ona hujayraning qobig'i ichida bo'lganida, ular kattalar o'simlikiga butunlay o'xshash bo'lib, o'z qobig'ini qo'yishadi.
Yosh xlorellalar o'sadi, etuklikka erishadi va sporogenez organiga aylanadi - idish bahs. Ona hujayrada 4-8 ta avtosporalar, qizi Xlorella paydo bo'ladi. Natijada, xlorellaning hayot aylanishi uchta tugun fazalari ketma-ketligi sifatida ifodalanadi: avtosport → vegetativ o'simlik → reproduktiv hujayra (konteyner) → avtosport va hokazo.
Shunday qilib, sporlar bilan ko'payish jarayonida oddiy hayot tsikli faqat uchta tugunli fazadan iborat ketma-ketlikka ega: 1 - o'simlikning boshlang'ich fazasi sifatida bir hujayrali rudiment, 2 - kattalar bir hujayrali yoki ko'p hujayrali organizm, 3 - ona (reproduktiv) hujayra. rudiment. Uchinchi fazadan keyin hayot yo'nalishi yana bir hujayrali rudiment fazasiga olib keladi.
Bunday oddiy hayot davrlari o'simliklar uchun xos emas. O'simlik guruhlarining aksariyati murakkab hayot davrlarini namoyish etadi. Ular odatda ikkita, ba'zan uchta oddiy tsiklni o'z ichiga oladi. Bundan tashqari, murakkab tsikllarda (jinsiy ko'payish paytida) 1-2 ta alohida bo'lishi kerak. gameta fazalari Va zigotalar .
Masalan, tabiatda gomosporali paporotnik ikki xil shaxslar shaklida ifodalanadi - paporotnikning o'zi va paporotnikning o'sishi. Fern prothallus (tuproqda zo'rg'a ko'rinadigan mayda yashil plitalar) yirik pinnate paporotnik shaxslarining bevosita avlodidir. U qisqa umr ko'radi, lekin bitta katta bargli shaxsning hayotini keltirib chiqarishga muvaffaq bo'ladi. Natijada, avlodlar almashinuvi mavjud: paporotnik → prothallus → paporotnik.
Spora bilan ko'payadigan paporotnik deyiladi sporofit (jinsiy avlod) va protallus gametalar orqali ko'payadi va deyiladi gametofit (jinsiy avlod). Gametofit va sporofit faqat shaxsni ko'paytirish usuli bilan aniqlanadi. Sporofit va gametofitning alohida mavjudligi mumkin emas va ular faqat avlodlar almashinadigan o'simliklarga tegishli.
Angiospermlarda urg'ochi gametofit odatda etti hujayragacha kamayadi, arxegoniyasi yo'q va embrion qop deb ataladi. Protallusga o'xshash embrion qopchasi mikroskopik jihatdan kichik va gulning chuqur qismida joylashgan.
Urug'li o'simliklarning erkak gametofiti mikrosporadan rivojlanadi va gulchang donasi (gulchang) bo'lib, ikki sperma hujayrasini hosil qilish uchun gulchang naychasiga aylanadi. Gullaydigan o'simlikning hayot aylanishi rasmda ko'rsatilgan. 12.

Guruch. 12. Gullaydigan o'simlikning hayot aylanishi
Jinsiy ko'payish partenogenetik va jinssiz ko'payish bilan almashinsa, hayot tsikllari sezilarli darajada murakkablashadi. Gaplo-diploid organizmlar mavjud bo'lib, ularda bir jins har doim faqat haplofazada, ikkinchisi esa diplo- va haplofazada bo'ladi. Bunday organizmlarga asal asalari kiradi (13-rasm).

Guruch. 13. Asalarilarning hayot aylanishi
Asalarilar koloniyasi bachadonining somatik hujayralari diploid bo'lib, gaplofaza faqat gametalar bilan ifodalanadi. Ishchi asalarilarda tuxumdonlar kichrayadi va uning hayot siklida gaplofaza bo'lmaydi. Dronlar urug'lanmagan tuxumdan partenogenetik tarzda rivojlanadi va xromosomalarning gaploid to'plamiga ega. Dronlarning gametogenezida meyozning mitoz bilan almashinishi tufayli ularning spermatozoidlari ham gaploid bo'lib chiqadi. Shuning uchun dronlar faqat gaplofazada mavjud.
Qo'ziqorinlar, ayniqsa, hayot aylanishlarida o'zgaruvchan (14-rasm). Ularning hayotiy tsiklida uchta yadro fazasi aniq belgilangan - haploid, diploid va dikarion.
Dikarion Ascomyces va Basidiomyces da uchraydi, ikkinchisida u tsiklning katta qismini tashkil qiladi.
Basidiomycesdagi gaploid holat o'tish davri bo'lib, diploid holat faqat zigota sifatida mavjud.
Zamburug'lar va suv o'tlarida haplofaza va diplofazaning davomiyligi nisbati o'zgaradi, shuning uchun hayot davrlarining turli oraliq variantlari kuzatiladi.

Guruch. 14. Zamburug'larning asosiy hayot sikllarining sxemasi
(yadro fazasidagi o'zgarishlar turli xil soyalar bilan belgilanadi,
o'qlar rivojlanish yo'nalishini ko'rsatadi)
Filogenez va ontogenez o'rtasidagi bog'liqlik 1. Ko'p hujayrali organizmlarning ontogenezi kattalashishi va tashkilotning murakkablashuvi yo'lidan boradi. 2. Organizmning ontogenezi ma'lum bir genetik dastur bo'yicha jinsiy etuklik va ko'payishning yakuniy maqsadi sari boradi. 3. Ko'p hujayralilikka o'tishi bilan ontogenez shakl jihatdan murakkablashadi va vaqt o'tishi bilan uzayadi. 4. Ontogenez evolyutsiyasining umumiy xususiyatlari quyidagilardan iborat: n ontogenezning avtonomizatsiyasi (dasturlanishi), n differensiallanish yo'nalishi, n metamorfozning yo'qolishi, epigenetik omillar ta'sirida rivojlanish dasturlari o'zgarishi ketma-ketligi. n
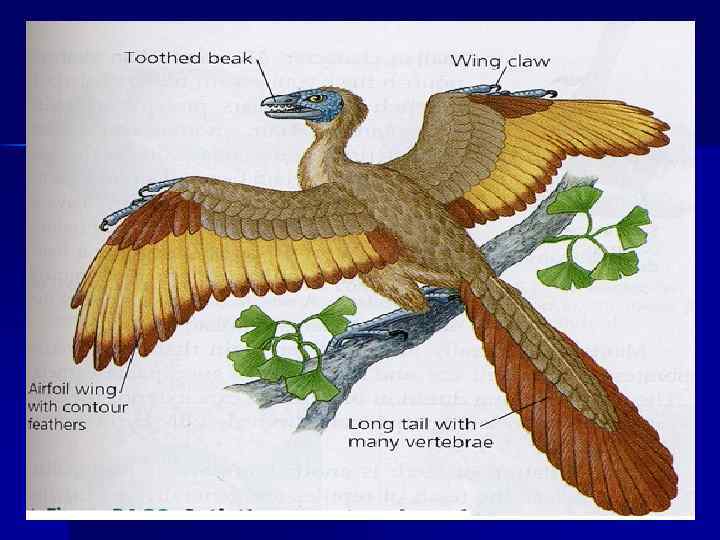
 Umurtqali hayvonlarning turli guruhlarida o'pkaning rivojlanishi: A - aksolotlda (Ambystoma); B - spadefoot spadefoot ichida (Pelobates); B - qurbaqada (V/o); G - kaltakesak (Lacerta), rivojlanishning I-III bosqichlari. Ikki marta soyali o'pkaning faqat nafas olish ta'sirida farqlanadigan qismlarini ko'rsatadi. Ko'rinib turibdiki, qurbaqa va kaltakesakda farqlanish faoliyat boshlanishidan oldin sodir bo'ladi (A. A. Mashkovtsev, 1936 yilga ko'ra).
Umurtqali hayvonlarning turli guruhlarida o'pkaning rivojlanishi: A - aksolotlda (Ambystoma); B - spadefoot spadefoot ichida (Pelobates); B - qurbaqada (V/o); G - kaltakesak (Lacerta), rivojlanishning I-III bosqichlari. Ikki marta soyali o'pkaning faqat nafas olish ta'sirida farqlanadigan qismlarini ko'rsatadi. Ko'rinib turibdiki, qurbaqa va kaltakesakda farqlanish faoliyat boshlanishidan oldin sodir bo'ladi (A. A. Mashkovtsev, 1936 yilga ko'ra).
 5. Ontogenezning turli bosqichlarida paydo bo'ladigan va yashashni oshiradigan moslashuvlar avlodlar o'rtasida tanlanish yo'li bilan saqlanib qoladi va organizmlar guruhining tarixiy rivojlanishining elementlari bo'ladi.
5. Ontogenezning turli bosqichlarida paydo bo'ladigan va yashashni oshiradigan moslashuvlar avlodlar o'rtasida tanlanish yo'li bilan saqlanib qoladi va organizmlar guruhining tarixiy rivojlanishining elementlari bo'ladi.
 Bir hujayrali evolyutsiya jarayonida ko'p hujayrali organizmlar ontogenezining murakkablashuvi sxemasi. Bir hujayrali Volvox tipidagi koloniyalar [hujayralar jinsiy (qora) va somatik bo'lib farqlanadi]. Gidra tipidagi ko'p hujayrali organizm (blastula va gastrula bosqichlari qo'shiladi). Birlamchi ikki tomonlama simmetrik hayvon (mezoderma qo'shiladi). Eng yuqori ikki tomonlama simmetrik hayvon.
Bir hujayrali evolyutsiya jarayonida ko'p hujayrali organizmlar ontogenezining murakkablashuvi sxemasi. Bir hujayrali Volvox tipidagi koloniyalar [hujayralar jinsiy (qora) va somatik bo'lib farqlanadi]. Gidra tipidagi ko'p hujayrali organizm (blastula va gastrula bosqichlari qo'shiladi). Birlamchi ikki tomonlama simmetrik hayvon (mezoderma qo'shiladi). Eng yuqori ikki tomonlama simmetrik hayvon.
 Germinal o'xshashlik qonuni. “Hayvonlarning ontogenezida birinchi navbatda yuqori taksonomik guruhlarning belgilari paydo bo'ladi va embrion rivojlanish jarayonida quyi taksonomik toifalarning xususiyatlari shakllanadi” (K.Baer) Bu genomning o'sha qismining konservatizmi bilan bog'liq. ontogenezdagi morfogenetik jarayonlar uchun javobgardir. Ammo filogenez jarayonida embriomutatsiyalar natijasida genlar oqimi yangilik, yangi belgilarga ega bo'ladi, ya'ni avlodlarning genotiplari ajdodlari genotiplaridan farq qiladi.
Germinal o'xshashlik qonuni. “Hayvonlarning ontogenezida birinchi navbatda yuqori taksonomik guruhlarning belgilari paydo bo'ladi va embrion rivojlanish jarayonida quyi taksonomik toifalarning xususiyatlari shakllanadi” (K.Baer) Bu genomning o'sha qismining konservatizmi bilan bog'liq. ontogenezdagi morfogenetik jarayonlar uchun javobgardir. Ammo filogenez jarayonida embriomutatsiyalar natijasida genlar oqimi yangilik, yangi belgilarga ega bo'ladi, ya'ni avlodlarning genotiplari ajdodlari genotiplaridan farq qiladi.

 Germinal o'xshashlik hodisasi. Rivojlanishning dastlabki bosqichidagi barcha umurtqali hayvonlarning embrionlari keyingi bosqichlarga qaraganda bir-biriga ko'proq o'xshaydi (E.Gekkel bo'yicha).
Germinal o'xshashlik hodisasi. Rivojlanishning dastlabki bosqichidagi barcha umurtqali hayvonlarning embrionlari keyingi bosqichlarga qaraganda bir-biriga ko'proq o'xshaydi (E.Gekkel bo'yicha).
 Ontogenez evolyutsiyasidagi irsiyat va oʻzgaruvchanlik rekapitulyatsiya, palingenez, senogenez, geteroxroniya (tezlanish, kechikish), geterotopiya va filembriogenez koʻrinishida namoyon boʻladi.
Ontogenez evolyutsiyasidagi irsiyat va oʻzgaruvchanlik rekapitulyatsiya, palingenez, senogenez, geteroxroniya (tezlanish, kechikish), geterotopiya va filembriogenez koʻrinishida namoyon boʻladi.
 F. Myuller evolyutsion qayta tashkil etish ikki xil bo'lishi mumkin deb hisoblagan. 1 Avlodlarning ontogenezi ajdodlarda tugagan bosqichdan keyin ham davom etishi mumkin. 2 Avlodlarning ontogenezi dastlabki yoki oraliq bosqichlarda ota-bobolari bosib oʻtgan yoʻldan chetga chiqishi mumkin. Biogenetik qonun Organizmning ontogenezi - ma'lum bir tur filogeniyasining qisqa va siqilgan takrorlanishi (rekapitulyatsiyasi) (E.Gekkel). A. N. Severtsov - 1. Ontogenez jarayonida katta yoshdagi ajdod shakllarining rivojlanish bosqichlari xususiyatlari takrorlanmasligi mumkin. 2. Ontogenez jarayonida yangi filogenetik yo'llar o'rnatilishi mumkin. Ontogenez jarayonida filogenetik ahamiyatga ega bo'lgan o'zgarishlarga filembriogenez deyiladi.
F. Myuller evolyutsion qayta tashkil etish ikki xil bo'lishi mumkin deb hisoblagan. 1 Avlodlarning ontogenezi ajdodlarda tugagan bosqichdan keyin ham davom etishi mumkin. 2 Avlodlarning ontogenezi dastlabki yoki oraliq bosqichlarda ota-bobolari bosib oʻtgan yoʻldan chetga chiqishi mumkin. Biogenetik qonun Organizmning ontogenezi - ma'lum bir tur filogeniyasining qisqa va siqilgan takrorlanishi (rekapitulyatsiyasi) (E.Gekkel). A. N. Severtsov - 1. Ontogenez jarayonida katta yoshdagi ajdod shakllarining rivojlanish bosqichlari xususiyatlari takrorlanmasligi mumkin. 2. Ontogenez jarayonida yangi filogenetik yo'llar o'rnatilishi mumkin. Ontogenez jarayonida filogenetik ahamiyatga ega bo'lgan o'zgarishlarga filembriogenez deyiladi.
 1. Anaboliya - morfogenezning yakuniy bosqichlarining kengayishi, ya'ni morfogenez jarayoni ajdodlarda morfogenez tugaydigan bosqichga etganidan keyin davom etadi. 2. Deviatsiya – og`ish, morfogenezning oraliq bosqichlarida rivojlanish yo`lining o`zgarishi. 3. Arxalaksis - organ shakllanishining dastlabki bosqichidagi o'zgarishlar, ya'ni uning rudimentlari. Morfogenez boshidanoq ajdodlardagidek davom etmaydi.
1. Anaboliya - morfogenezning yakuniy bosqichlarining kengayishi, ya'ni morfogenez jarayoni ajdodlarda morfogenez tugaydigan bosqichga etganidan keyin davom etadi. 2. Deviatsiya – og`ish, morfogenezning oraliq bosqichlarida rivojlanish yo`lining o`zgarishi. 3. Arxalaksis - organ shakllanishining dastlabki bosqichidagi o'zgarishlar, ya'ni uning rudimentlari. Morfogenez boshidanoq ajdodlardagidek davom etmaydi.
 Deviatsiya va archallaksis Suyak tarozi va sochlarning rivojlanishi: A - baliqlarning suyak tarozilari; B - sudralib yuruvchilarning shoxli tarozilari; B - sutemizuvchilarning sochlari. Yagona o'qlar - anaboliya, A dan B gacha - og'ish, B dan C gacha - ■ archallaxis. Soch paydo bo'lganda, boshlang'ich epidermal hujayralar guruhi tashqariga chiqmaydi, balki teriga tushadi; keyinchalik rudimentning butun rivojlanishi tarozilarning filogenetik rivojlanishini takrorlamaydi (A. N. Severtsov, 1939 yilga ko'ra).
Deviatsiya va archallaksis Suyak tarozi va sochlarning rivojlanishi: A - baliqlarning suyak tarozilari; B - sudralib yuruvchilarning shoxli tarozilari; B - sutemizuvchilarning sochlari. Yagona o'qlar - anaboliya, A dan B gacha - og'ish, B dan C gacha - ■ archallaxis. Soch paydo bo'lganda, boshlang'ich epidermal hujayralar guruhi tashqariga chiqmaydi, balki teriga tushadi; keyinchalik rudimentning butun rivojlanishi tarozilarning filogenetik rivojlanishini takrorlamaydi (A. N. Severtsov, 1939 yilga ko'ra).
 Filogenezda organlarning paydo bo'lishi, o'zgarishi va yo'q bo'lib ketishi organlarning filogenetik o'zgarishining negizida organlarning ko'p funksiyaliligi va ularning funktsiyalarini miqdoriy jihatdan o'zgartirish qobiliyati yotadi. 1. Organ qancha ko'p funktsiyani bajarsa, evolyutsiya jarayonida u qanchalik ko'p yo'nalishlarni o'zgartirishi mumkin. 2. Ko'p funktsiyalilik tizimlarning adaptiv transformatsiyasi imkoniyatini ta'minlaydi, 3. Funktsiyalarning mavjudligi yoki yo'qligi atrof-muhit o'zgarganda organlarni qayta qurish yo'nalishini belgilaydi. 4. Birlamchi va ikkilamchi funktsiyalar mavjud. 5. Funksiyalarning miqdoriy o‘zgarishlari irsiy heterojenlik va organ (to‘qima)ning bir jinsli morfofunksional birliklarining soni va o‘lchamidagi farqlardan kelib chiqadi.
Filogenezda organlarning paydo bo'lishi, o'zgarishi va yo'q bo'lib ketishi organlarning filogenetik o'zgarishining negizida organlarning ko'p funksiyaliligi va ularning funktsiyalarini miqdoriy jihatdan o'zgartirish qobiliyati yotadi. 1. Organ qancha ko'p funktsiyani bajarsa, evolyutsiya jarayonida u qanchalik ko'p yo'nalishlarni o'zgartirishi mumkin. 2. Ko'p funktsiyalilik tizimlarning adaptiv transformatsiyasi imkoniyatini ta'minlaydi, 3. Funktsiyalarning mavjudligi yoki yo'qligi atrof-muhit o'zgarganda organlarni qayta qurish yo'nalishini belgilaydi. 4. Birlamchi va ikkilamchi funktsiyalar mavjud. 5. Funksiyalarning miqdoriy o‘zgarishlari irsiy heterojenlik va organ (to‘qima)ning bir jinsli morfofunksional birliklarining soni va o‘lchamidagi farqlardan kelib chiqadi.
 Organlar va ularning funktsiyalarini o'zgartirishning asosiy usullari. 1. Ayrim organlarning asosiy funktsiyasini kuchaytirish ikki yo'l bilan amalga oshirilishi mumkin: a) organ tuzilishini o'zgartirish, b) yoki organdagi funktsional komponentlar sonini ko'paytirish orqali.
Organlar va ularning funktsiyalarini o'zgartirishning asosiy usullari. 1. Ayrim organlarning asosiy funktsiyasini kuchaytirish ikki yo'l bilan amalga oshirilishi mumkin: a) organ tuzilishini o'zgartirish, b) yoki organdagi funktsional komponentlar sonini ko'paytirish orqali.
 2. Asosiy funktsiyaning zaiflashishi. Organizm tuzilish va funktsiya nuqtai nazaridan optimal dizaynni ifodalaydi. Bu maqsadga muvofiqlik oziq-ovqat resurslarining surunkali etishmasligi va omon qolish uchun poyga natijasida yuzaga keladi. Amerikalik fiziologlar Teylor va Vaybel simmorfoz printsipini ishlab chiqdilar, hech qanday ishchi tuzilmaning kuchi maksimal yuk ostida tananing talab qiladigan darajadan oshadi. 3. Funktsiyalar sonini kamaytirish. Ixtisoslashgan organlar yoki tuzilmalar ba'zi funktsiyalarini yo'qotadi. 4. Funktsiyalar sonini ko'paytirish. A. alohida organlarning funktsiyalari sonining ko'payishi morfofunksional o'zgarishlar imkoniyatini oshiradi, b. Funktsiyalar soni ko'payganda, asosiysi, qoida tariqasida, o'zgarmaydi, lekin boshqalar tomonidan to'ldiriladi.
2. Asosiy funktsiyaning zaiflashishi. Organizm tuzilish va funktsiya nuqtai nazaridan optimal dizaynni ifodalaydi. Bu maqsadga muvofiqlik oziq-ovqat resurslarining surunkali etishmasligi va omon qolish uchun poyga natijasida yuzaga keladi. Amerikalik fiziologlar Teylor va Vaybel simmorfoz printsipini ishlab chiqdilar, hech qanday ishchi tuzilmaning kuchi maksimal yuk ostida tananing talab qiladigan darajadan oshadi. 3. Funktsiyalar sonini kamaytirish. Ixtisoslashgan organlar yoki tuzilmalar ba'zi funktsiyalarini yo'qotadi. 4. Funktsiyalar sonini ko'paytirish. A. alohida organlarning funktsiyalari sonining ko'payishi morfofunksional o'zgarishlar imkoniyatini oshiradi, b. Funktsiyalar soni ko'payganda, asosiysi, qoida tariqasida, o'zgarmaydi, lekin boshqalar tomonidan to'ldiriladi.
 5. Funktsiyalarning o'zgarishi odatda mavjudlik shartlari o'zgarganda sodir bo'ladi. Asosiysi o'z ahamiyatini yo'qotib qo'yishi mumkin, ikkinchi darajali funktsiyalardan biri esa asosiyning ahamiyatiga ega bo'lishi mumkin. 6. Organlarning polimerlanishi. 7. Organlarning oligomerizatsiyasi va funktsiyalarning konsentratsiyasi
5. Funktsiyalarning o'zgarishi odatda mavjudlik shartlari o'zgarganda sodir bo'ladi. Asosiysi o'z ahamiyatini yo'qotib qo'yishi mumkin, ikkinchi darajali funktsiyalardan biri esa asosiyning ahamiyatiga ega bo'lishi mumkin. 6. Organlarning polimerlanishi. 7. Organlarning oligomerizatsiyasi va funktsiyalarning konsentratsiyasi

 Evolyutsion munosabatlarning murakkabligi funktsiyalarni almashtirish, geterobatiya va kompensatsiya qilishda namoyon bo'ladi. Substitusiya - ontogenez va filogenez jarayonida organlar va funktsiyalarning boshqa organ bilan almashinishi A) Gomotopik almashtirish. B) Geterotopik almashtirish. Geterobatiyada alohida tizimlar va organlarning evolyutsiyasi turli tezliklarda sodir bo'ladi. Heterobatmiya organ tizimlarining mozaik evolyutsiyasining turli xil ixtisoslashuv tezligi. Kompensatsiya - ba'zi organlarning evolyutsiyasidagi kechikish boshqa organlarning tez o'zgarishi bilan qoplanishi mumkin.
Evolyutsion munosabatlarning murakkabligi funktsiyalarni almashtirish, geterobatiya va kompensatsiya qilishda namoyon bo'ladi. Substitusiya - ontogenez va filogenez jarayonida organlar va funktsiyalarning boshqa organ bilan almashinishi A) Gomotopik almashtirish. B) Geterotopik almashtirish. Geterobatiyada alohida tizimlar va organlarning evolyutsiyasi turli tezliklarda sodir bo'ladi. Heterobatmiya organ tizimlarining mozaik evolyutsiyasining turli xil ixtisoslashuv tezligi. Kompensatsiya - ba'zi organlarning evolyutsiyasidagi kechikish boshqa organlarning tez o'zgarishi bilan qoplanishi mumkin.
 Atavizmlar - bu ajdodlarida mavjud bo'lgan, ammo evolyutsiya jarayonida yo'qolgan ma'lum turdagi xususiyatlarning alohida organizmlarda paydo bo'lishi. Rudimentlar ajdod shakllarining gomologik organlariga nisbatan evolyutsiya jarayonida o'z funktsiyalarini amalda yo'qotgan rivojlanmagan organlardir. Organlarning rudimentatsiyasi. Evolyutsiya jarayonida funktsiyalar sonining kamayishi organning zaif rivojlanishiga olib kelishi mumkin. Rudimentlar odatda ba'zi funktsiyalarni bajaradilar, ular qoida tariqasida organning asl funktsiyasi bilan hech qanday aloqasi yo'q. Rudimentlar ko'pincha morfogenetik jarayonlarning faol bo'g'ini bo'lib, boshqa organlarning normal shakllanishini belgilaydi. Rudimentatsiya ikki yo'l bilan sodir bo'lishi mumkin: 1. Organning ontogenezi xuddi ajdodlarida bo'lgani kabi davom etadi, lekin u qaysidir bosqichda to'xtaydi. 2 Ontogenezda a'zoning og'rig'i ajdodlarga qaraganda kamroq yoki keyinchalik paydo bo'ladi, shuning uchun uning rivojlanishiga vaqt topa olmaydi.
Atavizmlar - bu ajdodlarida mavjud bo'lgan, ammo evolyutsiya jarayonida yo'qolgan ma'lum turdagi xususiyatlarning alohida organizmlarda paydo bo'lishi. Rudimentlar ajdod shakllarining gomologik organlariga nisbatan evolyutsiya jarayonida o'z funktsiyalarini amalda yo'qotgan rivojlanmagan organlardir. Organlarning rudimentatsiyasi. Evolyutsiya jarayonida funktsiyalar sonining kamayishi organning zaif rivojlanishiga olib kelishi mumkin. Rudimentlar odatda ba'zi funktsiyalarni bajaradilar, ular qoida tariqasida organning asl funktsiyasi bilan hech qanday aloqasi yo'q. Rudimentlar ko'pincha morfogenetik jarayonlarning faol bo'g'ini bo'lib, boshqa organlarning normal shakllanishini belgilaydi. Rudimentatsiya ikki yo'l bilan sodir bo'lishi mumkin: 1. Organning ontogenezi xuddi ajdodlarida bo'lgani kabi davom etadi, lekin u qaysidir bosqichda to'xtaydi. 2 Ontogenezda a'zoning og'rig'i ajdodlarga qaraganda kamroq yoki keyinchalik paydo bo'ladi, shuning uchun uning rivojlanishiga vaqt topa olmaydi.
 Vestigial organlarga misollar: A-pitonning orqa oyoq-qo'llari (Python regius); Kivining B qanoti (Apteryx australis); O‘ng kit (Eubalaena glacialis) tos kamarining B-elementlari (Avliyo Skovron bo‘yicha, 1965; A. A. Paramonov, 1978).
Vestigial organlarga misollar: A-pitonning orqa oyoq-qo'llari (Python regius); Kivining B qanoti (Apteryx australis); O‘ng kit (Eubalaena glacialis) tos kamarining B-elementlari (Avliyo Skovron bo‘yicha, 1965; A. A. Paramonov, 1978).
 Organlarning korrelyativ transformatsiyalari. Individual va tarixiy rivojlanishda ontogenezning yaxlitligi va barqarorligi morfogenez jarayonlarining korrelyatsiyasi va muvofiqlashuvi shaklida namoyon bo`ladi. Korrelyatsiyalar - rivojlanayotgan organizmning qismlari o'rtasidagi funktsional va tizimli munosabatlar. Genotipdagi genlarning o'zaro ta'siri va bog'lanishiga asoslangan genomik korrelyatsiya. Morfogenetik korrelyatsiyalar embriogenez jarayonida hujayralar yoki tana qismlarining bir-biri bilan o'zaro ta'siriga asoslanadi. Ergontik korrelyatsiyalar aniq tuzilmalar o'rtasida funktsional bog'liqlikni o'rnatadi. Muvofiqlashtirish. Filogenezda organlardagi bog'liq o'zgarishlar. Organlarning fazoviy aloqalarini topografik muvofiqlashtirish. Funktsional ravishda bir-biriga bog'langan organlar va ularning tizimlarining filogenez jarayonidagi o'zgarishlarni dinamik muvofiqlashtirish. Bir-biri bilan bevosita bog'liq bo'lmagan organlardagi evolyutsion o'zgarishlarni biologik muvofiqlashtirish. Tanlov hayotni ta'minlash uchun ularning muvofiqlashtirilgan o'zgarishiga qarab harakat qiladi
Organlarning korrelyativ transformatsiyalari. Individual va tarixiy rivojlanishda ontogenezning yaxlitligi va barqarorligi morfogenez jarayonlarining korrelyatsiyasi va muvofiqlashuvi shaklida namoyon bo`ladi. Korrelyatsiyalar - rivojlanayotgan organizmning qismlari o'rtasidagi funktsional va tizimli munosabatlar. Genotipdagi genlarning o'zaro ta'siri va bog'lanishiga asoslangan genomik korrelyatsiya. Morfogenetik korrelyatsiyalar embriogenez jarayonida hujayralar yoki tana qismlarining bir-biri bilan o'zaro ta'siriga asoslanadi. Ergontik korrelyatsiyalar aniq tuzilmalar o'rtasida funktsional bog'liqlikni o'rnatadi. Muvofiqlashtirish. Filogenezda organlardagi bog'liq o'zgarishlar. Organlarning fazoviy aloqalarini topografik muvofiqlashtirish. Funktsional ravishda bir-biriga bog'langan organlar va ularning tizimlarining filogenez jarayonidagi o'zgarishlarni dinamik muvofiqlashtirish. Bir-biri bilan bevosita bog'liq bo'lmagan organlardagi evolyutsion o'zgarishlarni biologik muvofiqlashtirish. Tanlov hayotni ta'minlash uchun ularning muvofiqlashtirilgan o'zgarishiga qarab harakat qiladi
 Tug'ma nuqsonlar Rivojlanish nuqsonlari - tug'ilishdan oldin sodir bo'lgan, darhol aniqlanadigan yoki tug'ilgandan keyin bir muncha vaqt o'tgach, organlar faoliyatining buzilishiga olib keladigan tizimli anomaliyalar. I. Sabablariga koʻra tugʻma nuqsonlar 1. Irsiy (gen yoki xromosomalarning oʻzgarishi natijasida kelib chiqadigan, biokimyoviy, hujayra, toʻqima va aʼzo va organizm jarayonlarining buzilishiga olib keladigan) ga boʻlinadi. 2. Ekzogen (teratogen muhit omillari ta'sirida paydo bo'ladi). Teratogenlar mutatsiyalar bilan bir xil jarayonlarga ta'sir qilganligi sababli, ekzogen va genetik nuqsonlarning fenotipik namoyon bo'lishi juda o'xshash bo'lishi mumkin, bu fenokopiya atamasi bilan belgilanadi. 3. Ko'p omilli nuqsonlar ekzogen va irsiy omillar ta'sirida yuzaga keladi.
Tug'ma nuqsonlar Rivojlanish nuqsonlari - tug'ilishdan oldin sodir bo'lgan, darhol aniqlanadigan yoki tug'ilgandan keyin bir muncha vaqt o'tgach, organlar faoliyatining buzilishiga olib keladigan tizimli anomaliyalar. I. Sabablariga koʻra tugʻma nuqsonlar 1. Irsiy (gen yoki xromosomalarning oʻzgarishi natijasida kelib chiqadigan, biokimyoviy, hujayra, toʻqima va aʼzo va organizm jarayonlarining buzilishiga olib keladigan) ga boʻlinadi. 2. Ekzogen (teratogen muhit omillari ta'sirida paydo bo'ladi). Teratogenlar mutatsiyalar bilan bir xil jarayonlarga ta'sir qilganligi sababli, ekzogen va genetik nuqsonlarning fenotipik namoyon bo'lishi juda o'xshash bo'lishi mumkin, bu fenokopiya atamasi bilan belgilanadi. 3. Ko'p omilli nuqsonlar ekzogen va irsiy omillar ta'sirida yuzaga keladi.
 II. Prenatal ontogenez bosqichiga qarab: 1. Gametopatiyalar (zigota bosqichidagi rivojlanish buzilishlari) 2. Blastopatiyalar (blastula bosqichidagi rivojlanish buzilishlari) 3. Embriopatiyalar (15 kundan 8 haftagacha bo'lgan davrdagi buzilishlar) 4. Fetopatiyalar (dizorlar). 10 haftadan keyin sodir bo'lgan ) Embrion morfogenezining buzilishi (3-10 hafta) ko'pincha buzilishlar natijasida nuqsonlarga olib keladi: ko'payish (organlarning gipoplaziyasi va aplaziyasi), migratsiya (heterotopiya), differentsiatsiya (embrion tuzilmalarining barqarorligi, aplaziya). organ yoki uning bir qismi), adezyon va hujayra o'limi (fuziya disrafizmi).
II. Prenatal ontogenez bosqichiga qarab: 1. Gametopatiyalar (zigota bosqichidagi rivojlanish buzilishlari) 2. Blastopatiyalar (blastula bosqichidagi rivojlanish buzilishlari) 3. Embriopatiyalar (15 kundan 8 haftagacha bo'lgan davrdagi buzilishlar) 4. Fetopatiyalar (dizorlar). 10 haftadan keyin sodir bo'lgan ) Embrion morfogenezining buzilishi (3-10 hafta) ko'pincha buzilishlar natijasida nuqsonlarga olib keladi: ko'payish (organlarning gipoplaziyasi va aplaziyasi), migratsiya (heterotopiya), differentsiatsiya (embrion tuzilmalarining barqarorligi, aplaziya). organ yoki uning bir qismi), adezyon va hujayra o'limi (fuziya disrafizmi).

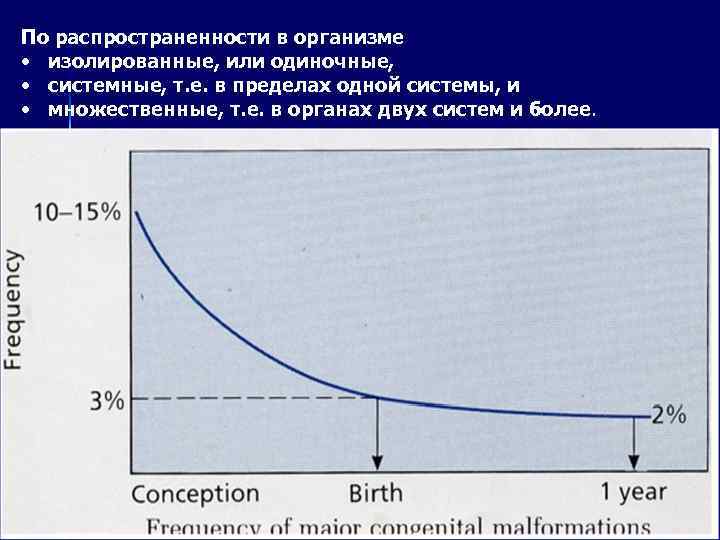 Tanadagi tarqalishiga ko'ra, izolyatsiya qilingan yoki bitta, tizimli, ya'ni bir tizim ichida va ko'p, ya'ni ikki yoki undan ortiq tizim organlarida.
Tanadagi tarqalishiga ko'ra, izolyatsiya qilingan yoki bitta, tizimli, ya'ni bir tizim ichida va ko'p, ya'ni ikki yoki undan ortiq tizim organlarida.
 Tizimli tug'ma nuqsonlar etiologiyasi Ideopatik 60% Multifaktorli 20% Monogen 7. 5% Xromosoma 6% Onalik kasalliklari 3% Bachadon ichidagi infektsiyalar 2% Dorilar, nurlanish, alkogol va boshqalar 1. 5%
Tizimli tug'ma nuqsonlar etiologiyasi Ideopatik 60% Multifaktorli 20% Monogen 7. 5% Xromosoma 6% Onalik kasalliklari 3% Bachadon ichidagi infektsiyalar 2% Dorilar, nurlanish, alkogol va boshqalar 1. 5%
 Filogenetik ahamiyatiga ko'ra malformatsiyalar filogenetik va filogenetik bo'lmaganlarga bo'linadi. Filogenetik jihatdan aniqlangan nuqsonlar chordatlar va umurtqalilar kenja tipidagi hayvonlarning organlariga o'xshaydi (ajdodlar yoki atavistik nuqsonlar). Ular odamlarning boshqa umurtqali hayvonlar bilan genetik aloqasini ko'rsatadi, shuningdek, nuqsonlarning paydo bo'lish mexanizmlarini tushunishga yordam beradi. Atavizmlarning paydo bo'lishining etakchi mexanizmlari, ehtimol, morfogenez tezligini va organlarni qisqartirishga qaratilgan jarayonlarni boshlashni nazorat qiluvchi tartibga soluvchi genlarning mutatsiyalari. Eng keng tarqalgan atavizmlar: Morfogenez bosqichlarida organlarning rivojlanmaganligi, ular ajdodlar holatini takrorlagan. Embrion tuzilmalarining barqarorligi, shuningdek, ajdodlarga xos bo'lgan morfologiyani takrorlaydi. Ontogenezda organlar harakatining buzilishi.
Filogenetik ahamiyatiga ko'ra malformatsiyalar filogenetik va filogenetik bo'lmaganlarga bo'linadi. Filogenetik jihatdan aniqlangan nuqsonlar chordatlar va umurtqalilar kenja tipidagi hayvonlarning organlariga o'xshaydi (ajdodlar yoki atavistik nuqsonlar). Ular odamlarning boshqa umurtqali hayvonlar bilan genetik aloqasini ko'rsatadi, shuningdek, nuqsonlarning paydo bo'lish mexanizmlarini tushunishga yordam beradi. Atavizmlarning paydo bo'lishining etakchi mexanizmlari, ehtimol, morfogenez tezligini va organlarni qisqartirishga qaratilgan jarayonlarni boshlashni nazorat qiluvchi tartibga soluvchi genlarning mutatsiyalari. Eng keng tarqalgan atavizmlar: Morfogenez bosqichlarida organlarning rivojlanmaganligi, ular ajdodlar holatini takrorlagan. Embrion tuzilmalarining barqarorligi, shuningdek, ajdodlarga xos bo'lgan morfologiyani takrorlaydi. Ontogenezda organlar harakatining buzilishi.




Har xil turdagi individlarning ontogenezi farqlanish davomiyligi, tezligi va tabiati jihatidan bir xil emas (pastga qarang). Odatda proembrional, embrion va postembrional davrlarga bo'linadi. Hayvonlarda embrion davri odatda differensiatsiyaga boy, o'simliklarda esa postembrional davr boy. Ushbu ontogenez davrlarining har birini ketma-ket sifat bosqichlariga bo'lish mumkin. Ontogenez to'g'ridan-to'g'ri rivojlanish yoki metamorfoz orqali rivojlanish bilan tavsiflanishi mumkin.
Turli guruhlardagi ontogenezning xususiyatlari. Tirik tabiatda individuallikning namoyon bo'lish shakllari xilma-xil bo'lib, prokariotlar, zamburug'lar, o'simliklar va hayvonlarning turli vakillarida ontogenez jarayoni mazmunan teng emas.
Guruch. 14.1. Evolyutsiya jarayonida ko'p hujayrali organizmlar ontogenezining ketma-ket murakkablashuvi sxemasi. A - erkin yashovchi bir hujayrali organizmlarning ko'payishi; B - bir hujayrali Volvox tipidagi koloniyaning ontogenezi [hujayralar jinsiy (qora) va somatik bo'linadi]; B - gidra tipidagi ko'p hujayrali organizmning ontogenezi (blastula va gastrula bosqichlari qo'shiladi); D - birlamchi ikki tomonlama simmetrik hayvonning ontogenezi (mezoderma qo'shiladi); D - eng yuqori ikki tomonlama simmetrik hayvonning ontogenezi (A.N. Severtsov bo'yicha, 1935 yil)
Ko'p hujayralilikka (Metazoa) o'tishi bilan ontogenez shakl jihatidan murakkablashadi va vaqt o'tishi bilan uzayadi (14.1-rasm), lekin ontogenez evolyutsiyasi jarayonida rivojlanishning soddalashtirilgan holatlari ham kuzatiladi, bu ko'proq hujayralarning paydo bo'lishi bilan bog'liq. irsiy axborotni amalga oshirishning ilg'or usullari. Evolyutsiya jarayonida o'simliklar va hayvonlar murakkab rivojlanish davrlarini boshdan kechiradilar, ularning har bir bosqichi muayyan atrof-muhit sharoitlariga moslashadi. Ba'zan evolyutsiya jarayonida hayot davrlarining ikkilamchi soddalashuvi sodir bo'ladi.
Hayotiy tsiklning soddalashtirilishi bilan ontogenetik rivojlanishning butun jarayoni sifat jihatidan o'zgaradi. Hayotiy tsiklni soddalashtirishning oqibatlaridan biri rivojlanishning gaploid bosqichidan diploid fazaga va metamorfoz bilan rivojlanishdan (masalan, amfibiyalarda) to'g'ridan-to'g'ri rivojlanishga (sudraluvchilar va boshqa yuqori umurtqali hayvonlarda) o'tishdir. To'g'ridan-to'g'ri rivojlanishi bilan yangi tug'ilgan hayvon kattalar mavjudotining barcha asosiy tashkiliy xususiyatlariga ega. Metamorfoz bilan rivojlanish bir qator lichinka bosqichlaridan o'tadi; Tuxumdan lichinka paydo bo'ladi va murakkab o'zgarish orqali kattalar hayvoniga xos xususiyatlarni oladi. Rivojlanishdan metamorfoz orqali to'g'ridan-to'g'ri rivojlanishga o'tish Yerdagi hayot evolyutsiyasining so'nggi bosqichlarining eng muhim natijalaridan biridir.
Daraxtlar, butalar va ko'p yillik o'tlardagi individning murakkab bo'linishiga qaramasdan, ontogenezning tashkiliy darajasi bo'yicha ular bir yillik, ikki yillik va efemer gulli o'simliklardan pastroqdir. Ikkinchisida ontogenez ma'lum miqdordagi organlarning hayotiy faoliyatini qat'iy muvofiqlashtirish bilan davom etadi. Ularning ontogenezidagi differentsiatsiya va morfogenez jarayonlari tabiatda "portlovchi" dir.
O'simliklarda ontogenez tartibga solish tizimining zaif rivojlanishi tufayli katta labillik bilan tavsiflanadi (pastga qarang). Umuman o'simliklardagi ontogenez hayvonlarnikiga qaraganda ko'proq atrof-muhit sharoitlariga bog'liq.
Turli organizmlarda ontogenezning umumiy xususiyatlari uning dasturlanishi, differensiallanish yo'nalishi, atrof-muhit omillari (epigenetik omillar) ta'sirida rivojlanish dasturlaridagi o'zgarishlar ketma-ketligidir.
Organizmlarning turli guruhlaridagi ontogenezning xilma-xilligi (hatto bir xil tur vakillari orasida ham) differensiatsiya va hayot davrlarini barqarorlashtirishda atrof-muhit omillarining alohida rolini ko'rsatadi. Tanlov yaxlit ontogenez bo'ylab sodir bo'lsa-da, uning alohida bosqichlari butun dasturni amalga oshirish va avlodlar o'rtasidagi axborot oqimi uchun zarur shartlar bo'lib xizmat qiladi.
Turli shohliklar, tiplar, sinflar vakillarida ontogenez differensiatsiya ko'lami bilan ham farqlanadi. Bir hujayrali organizmlarda differensiatsiya jarayonlarining murakkabligi jihatidan primitivdir. O'simliklarda differentsiatsiya jarayonlari uzaytiriladi va embrion rivojlanish davri bilan cheklanmaydi (o'simliklarda metamerik organlarning shakllanishi ontogenez davomida sodir bo'ladi). Hayvonlarda differensiatsiya va organlar hosil bo'lish jarayonlari birinchi navbatda embrion davri bilan chegaralanadi. O'simliklardagi gistogenez va morfogenez jarayonlari hayvonlarnikiga qaraganda kamroq murakkab va kamroq organ va tuzilmalarni o'z ichiga oladi.
Ontogenezning davomiyligi. Har xil turdagi, sinflar, tartiblar vakillarida ontogenezning davomiyligi muhim tur xususiyati hisoblanadi. Tabiiy o'limning boshlanishi bilan umr ko'rish davomiyligining cheklanishi, hatto qulay tashqi sharoitlar mavjud bo'lganda ham, evolyutsiyaning muhim natijasi bo'lib, avlodlar almashinuviga imkon beradi. Bir hujayrali organizmlarda ontogenez qiz hujayralarining shakllanishi bilan tugaydi, o'lim morfologik jihatdan aniqlanmaydi (va ular ma'lum ma'noda o'lmas). Zamburug'lar va o'simliklarda turli organlarning qarishi notekis sodir bo'ladi. Qo'ziqorinlarda "mitseliy" ning o'zi uzoq vaqt davomida substratda yashaydi (o'tloqli asal qo'ziqorinida (Marasmius oreades) - 500 yilgacha!). Boshqa tomondan, qo'ziqorinlar orasida haftalar va oylar davomida yashaydigan efemer organizmlar mavjud (Clavaria gyromitra). Jadvalda 14.1-jadvalda bir qator o'simliklarning umr ko'rish davomiyligi bo'yicha ba'zi ma'lumotlar keltirilgan. O'simliklar ham, xuddi hayvonlar kabi, insonning umri davomida juda xilma-xildir.
| 14.1-jadval. Ayrim turlarning ontogenez davomiyligi | |
| Turlari | Ontogenezning davomiyligi |
| 1. Yadrodan oldingi qirollik | |
| Cyanei | Bir necha soat |
| II. qo'ziqorinlar shohligi | |
| Penicillium notatum | Bir necha hafta |
| Polipore (Fomes fomentarius) | 25 yilgacha |
| Oq qo'ziqorin (Botulus botulus) | Bir necha yillar |
| III. o'simliklar shohligi | |
| Kesish (Arabidopsis thaliana) | 60-70 kun |
| Bug'doy (Triticum bulgare) | Taxminan 1 yil |
| Uzum (Vitis vinifera) | 80-100 yil |
| Olma daraxti (Malus domestica) | 200 yil |
| Yong'oq (Juglans regia) | 300-400 yil |
| Linden (Tilia grandifolia) | 1000 yil |
| Eman (Quercus robur) | 1200 yil |
| Sarv (Cupressus fastigiata) | 3000 yil |
| Mamont daraxti (Sequoia gigantea) | 5000 yil |
| IV. hayvonlar shohligi | |
| Keng tasmasimon qurt (Diphyllobothrium latum) | 29 yoshgacha |
| Chumoli (Formica fusca) | 7 yilgacha |
| Asal ari (Apis mellifera) | 5 yilgacha |
| Dengiz kirpisi (Ehinus esculentus) | 8 yilgacha |
| Com (Silurus glanis) | 60 yoshgacha |
| Gobi (Aphya pellucida) | 1 yil |
| Oddiy qurbaqa (Bufo bufo) | 36 yoshgacha |
| Toshbaqa (Testudo sumelri) | 150 yilgacha |
| Oddiy burgut boyo'g'li (Bubo bubo) | 68 yoshgacha |
| Qoya kaptar (Columba livid) | 30 yoshgacha |
| Afrika fili (Elephas maximus) | 60 yoshgacha |
| Gibbon (Hylobates lar) | 32 yoshgacha |
Ontogenez- shaxsning individual rivojlanishi, zigota paydo bo'lgan paytdan to o'limgacha bo'lgan hayot tsikli davomida tabiiy ravishda sodir bo'ladigan o'zaro bog'liq o'zgarishlarning umumiyligi.
Jinsiy yo'l bilan ko'payadigan ko'p hujayrali hayvonlarda ontogenez bo'linadi embrion(zigota hosil bo'lishidan tug'ilish yoki tuxum membranalaridan chiqishgacha) va postembrionik(tuxum membranalaridan chiqish yoki organizmning tug'ilishidan o'limigacha) davrlar. Erkak va ayol jinsiy hujayralari - gametalarning birlashishi natijasida zigota hosil bo'ladi. Gametalar jinsiy bezlarda erkak yoki urg'ochi organizmga qarab hosil bo'ladi. Gametalarning rivojlanish jarayoni deyiladi gametogenez. Sperma hosil bo'lish jarayoni deyiladi spermatogenez, va tuxum shakllanishi hisoblanadi oogenez.
Spermatogenez
1 - ko'payish bosqichi; 2 - o'sish bosqichi; 3 - etuklik bosqichi; 4 - shakllanish bosqichi.
Spermatogenez moyaklar ichida sodir bo'lib, to'rt bosqichga bo'linadi: 1) ko'payish, 2) o'sish, 3) etilish, 4) hosil bo'lish. Ko'payish bosqichida diploid spermatozoidlar mitoz yo'li bilan qayta-qayta bo'linadi. Olingan spermatogoniyalarning ba'zilari takroriy mitotik bo'linishlarga duchor bo'lishi mumkin, natijada bir xil sperma hujayralari paydo bo'ladi. Boshqa qismi bo'linishni to'xtatadi va hajmi kattalashib, spermatogenezning keyingi bosqichiga - o'sish bosqichiga kiradi.
O'sish bosqichi meiozning 1 interfazasiga to'g'ri keladi, ya'ni. Bu jarayonda hujayralar meiozga tayyorlanadi. O'sish fazasining asosiy hodisasi DNK replikatsiyasidir. Yetilish fazasida hujayralar meioz yo‘li bilan bo‘linadi; birinchi meyotik bo'linish paytida ular deyiladi 1-tartib spermatotsitlar, ikkinchi davrda - 2-tartib spermatotsitlar. Bitta birinchi tartibli spermatotsitdan to'rtta gaploid spermatidlar paydo bo'ladi. Shakllanish fazasi dastlab sharsimon spermatidlar bir qator murakkab o'zgarishlarga uchrab, buning natijasida spermatozoidlar hosil bo'lishi bilan tavsiflanadi. Unda yadro va sitoplazmaning barcha elementlari ishtirok etadi.
Odamlarda spermatogenez balog'at yoshida boshlanadi; Sperma shakllanishi davri uch oy, ya'ni. sperma har uch oyda yangilanadi. Spermatogenez millionlab hujayralarda doimiy va sinxron tarzda sodir bo'ladi.

1 - "bosh"; 2 - "bo'yin"; 3 - o'rta qism; 4 - flagellum; 5 - akrozoma; 6 - yadro; 7 - sentriolalar; 8 - mitoxondriya.
Sutemizuvchilar spermasi uzun ipga o'xshaydi. Odam spermatozoidining uzunligi 50-60 mikron. Spermatozoidning tuzilishini "bosh", "bo'yin", oraliq bo'lim va quyruqga bo'lish mumkin. Boshda yadro va akrozoma mavjud. Yadroda gaploid xromosomalar to'plami mavjud. Akrozoma - bu tuxum membranalarini eritish uchun ishlatiladigan fermentlarni o'z ichiga olgan membrana organellasi. Bo'yin qismida ikkita sentriola, oraliq qismida mitoxondriya mavjud. Quyruq bitta, ba'zi turlarda - ikki yoki undan ortiq flagella bilan ifodalanadi. Flagellum harakat organellasi bo'lib, tuzilishi jihatidan oddiy hayvonlarning flagella va kirpiklariga o'xshaydi. Flagella harakati uchun ATP ning makroergik aloqalari energiyasi ishlatiladi, ATP sintezi mitoxondriyalarda sodir bo'ladi.
Spermatozoid 1677 yilda A. Leuvenguk tomonidan kashf etilgan.
Oogenez
U tuxumdonlarda amalga oshiriladi va uch fazaga bo'linadi: 1) ko'payish, 2) o'sish, 3) kamolot.
Reproduktiv fazada diploid oogoniyalar mitoz yo'li bilan qayta-qayta bo'linadi. O'sish bosqichi meiozning 1 interfazasiga to'g'ri keladi, ya'ni. uning davomida hujayralar meiozga tayyorlanadi: ozuqa moddalarining to'planishi tufayli hujayralar sezilarli darajada oshadi. O'sish fazasining asosiy hodisasi DNK replikatsiyasidir. Yetilish bosqichida hujayralar meioz yo'li bilan bo'linadi. Birinchi meyotik bo'linish paytida ular chaqiriladi 1-tartibli oositlar. Birinchi meiotik bo'linish natijasida ikkita qiz hujayra paydo bo'ladi: birinchisi deb ataladigan kichik qutb tanasi, va kattaroq - 2-tartibli oosit. Ikkinchi meyotik boʻlinish jarayonida 2-tartibli oosit boʻlinib tuxum va ikkinchi qutb tanasini, birinchi qutb tanasi boʻlinib uchinchi va toʻrtinchi qutb tanachalarini hosil qiladi. Shunday qilib, meioz natijasida 1-tartibdagi bir oositdan bitta oosit va uchta qutb tanasi hosil bo'ladi.

1—koʻpayish bosqichi; 2 - o'sish bosqichi; 3 - etuklik bosqichi.
Balog'atga etganidan keyingina sodir bo'ladigan sperma shakllanishidan farqli o'laroq, odamlarda tuxum shakllanishi jarayoni embrional davrda boshlanadi va vaqti-vaqti bilan davom etadi. Embrionda ko'payish va o'sish fazalari to'liq amalga oshiriladi va kamolot davri boshlanadi. Qiz tug'ilganda, uning tuxumdonlarida yuz minglab birinchi tartibli oositlar mavjud bo'lib, ular to'xtab qolgan, meyozning 1-fazasining diploten bosqichida "muzlatilgan" - Oogenezning birinchi bloki.
Jinsiy balog'at davrida meyoz qayta boshlanadi: taxminan har oy jinsiy gormonlar ta'sirida oositlardan biri (kamdan-kam hollarda ikkita) meiozning 2 metafazasiga etadi - Oogenezning ikkinchi bloki. Meioz faqat urug'lanish sodir bo'lgan taqdirdagina yakunlanishi mumkin; agar urug'lantirish sodir bo'lmasa, ikkinchi tartibli oosit o'ladi va tanadan chiqariladi.
Tuxumlarning tuzilishi
Tuxumlarning shakli odatda yumaloq bo'ladi. Tuxumlarning o'lchamlari juda xilma-xildir - bir necha o'nlab mikrometrdan bir necha santimetrgacha (odam tuxumi taxminan 120 mikron). Tuxumlarning strukturaviy xususiyatlariga quyidagilar kiradi: plazma membranasining tepasida joylashgan membranalarning mavjudligi va sitoplazmada ko'p yoki kamroq zaxira ozuqa moddalarining mavjudligi.

1 - pronukleus 2 meta-fazada; 2 - porloq qobiq; 3— radiatsion qobiq; 4 - birinchi qutb tanasi.
Ko'pgina hayvonlarda tuxum sitoplazmatik membrananing tepasida joylashgan qo'shimcha membranalarga ega. Kelib chiqishiga qarab, ular birlamchi, ikkilamchi va uchinchi darajali qobiqlarga bo'linadi. Birlamchi qobiqlar oosit tomonidan ajratilgan moddalardan hosil bo'ladi. Tuxumning sitoplazmatik membranasi bilan aloqa qilishda qatlam hosil bo'ladi. U himoya funktsiyasini bajaradi, spermatozoidlarning kirib borishining turlarga xosligini ta'minlaydi, ya'ni. boshqa turdagi spermatozoidlarning tuxumga kirishiga to'sqinlik qiladi. Sutemizuvchilarda bu membrana pellucida deb ataladi. Ikkilamchi qobiqlar tuxumdon follikulyar hujayralarining sekretsiyasi bilan hosil bo'ladi va barcha tuxumlarda mavjud emas. Hasharot tuxumlarining ikkilamchi qobig'ida sperma tuxumga kirib boradigan kanal - mikropila mavjud. Uchinchi darajali qobiqlar tuxum yo'llarining maxsus bezlari faoliyati tufayli hosil bo'ladi. Masalan, maxsus bezlar sekretsiyasidan qushlar va sudralib yuruvchilarda oqsil, qobiq osti pergament, qobiq va qobiq ustki membranalar hosil bo'ladi.
Hayvon tuxumlarida ikkilamchi va uchinchi darajali membranalar hosil bo'lib, ularning embrionlari tashqi muhitda rivojlanadi. Sutemizuvchilar bachadon ichi rivojlanishiga to'g'ri kelganligi sababli, ularning tuxumlari faqat birlamchi membranaga ega bo'lib, uning tepasida toj radiata, tuxumga ozuqa moddalarini etkazib beradigan follikulyar hujayralar qatlami joylashgan.
Tuxumlarda ozuqa moddalarining zaxirasi to'planadi, ular deyiladi sarig'i. U yog'lar, uglevodlar, RNK, minerallar, oqsillarni o'z ichiga oladi, ularning asosiy qismini lipoproteinlar va glikoproteinlar tashkil qiladi. Sarig'i sitoplazmada bo'ladi sarig'i granulalari. Tuxumda to'plangan oziq moddalar miqdori embrionning rivojlanishi sharoitlariga bog'liq. Agar tuxumning rivojlanishi onaning tanasidan tashqarida sodir bo'lsa va yirik hayvonlarning shakllanishiga olib keladigan bo'lsa, unda sarig'i tuxum hajmining 95% dan ko'prog'ini tashkil qilishi mumkin. Onaning tanasida rivojlanadigan sutemizuvchilar tuxumlari oz miqdorda sarig'ni o'z ichiga oladi - 5% dan kam, chunki embrionlar rivojlanish uchun zarur bo'lgan ozuqa moddalarini onadan oladi.

1 - aletital; 2 - izol-sital; 3 - o'rtacha telolesital; 4 - keskin telolesital.
Tarkibidagi sarig'i miqdoriga qarab, quyidagilar ajralib turadi: tuxum turlari: aletital(sarig'i yo'q yoki oz miqdorda sarig' qo'shilishi - sutemizuvchilar, yassi qurtlar); izolesital(tekis taqsimlangan sarig'i bilan - lancelet, dengiz kirpi); o'rtacha telolesital(notekis taqsimlangan sarig'i bilan - baliq, amfibiyalar); keskin telolesital(sarig'i eng katta qismini egallaydi va hayvon qutbidagi sitoplazmaning faqat kichik bir qismi - qushlar undan ozoddir).
Oziq moddalarning to'planishi tufayli tuxumlar polaritni rivojlantiradi. Qarama-qarshi qutblar deyiladi vegetativ Va hayvoniy. Polarizatsiya hujayradagi yadroning joylashishining o'zgarishida (u hayvon qutbi tomon siljiydi), shuningdek sitoplazmatik qo'shimchalarning tarqalishida (ko'p tuxumlarda sarig'i miqdori hayvondan ko'payadi) namoyon bo'ladi. vegetativ qutbga).
Inson tuxumini 1827 yilda K.M. Baer.
Urug'lantirish
Erkak va urg'ochi jinsiy hujayralarning qo'shilib, zigota hosil bo'lishiga olib keladigan va yangi organizmni keltirib chiqaradigan jarayon deyiladi. urug'lantirish. Urug'lantirish jarayonining o'zi sperma va tuxum o'rtasidagi aloqa paytidan boshlanadi. Bunday aloqa paytida akrosomal o'simtaning plazma membranasi va akrosomal pufakcha membranasining qo'shni qismi eriydi, gialuronidaza fermenti va akrozoma tarkibidagi boshqa biologik faol moddalar ajralib chiqadi va tuxum membranasining bir qismini eritadi. . Ko'pincha sperma tuxum ichiga to'liq tortiladi, ba'zida flagellum tashqarida qoladi va tashlanadi. Spermatozoid tuxumga kirgan paytdan boshlab gametalar o'z faoliyatini to'xtatadi, chunki ular bitta hujayra - zigota hosil qiladi. Sperma yadrosi shishadi, uning xromatini bo'shashadi, yadro membranasi eriydi va u erkak pronukleus. Bu urug'lanish tufayli qayta boshlangan tuxum yadrosining ikkinchi meiotik bo'linishi tugashi bilan bir vaqtda sodir bo'ladi. Asta-sekin tuxum yadrosi aylanadi ayol pronukleus. Pronukleuslar tuxumning markaziga o'tadi, DNK replikatsiyasi sodir bo'ladi va ular birlashgandan so'ng xromosomalar to'plami va zigota DNKsi "2" ga aylanadi. n 4c" Pronukleuslarning birlashishi ifodalaydi haqiqiy urug'lantirish. Shunday qilib, urug'lantirish diploid yadroli zigota hosil bo'lishi bilan tugaydi.
Urug'lantirish qaytarib bo'lmaydigan jarayondir, ya'ni urug'lantirilgan tuxum qayta urug'lantirilmaydi.
Jinsiy ko'payishda ishtirok etadigan shaxslar soniga qarab quyidagilar ajralib turadi: o'zaro urug'lantirish- urug'lanish, unda turli organizmlar tomonidan hosil bo'lgan gametalar ishtirok etadi; o'z-o'zini urug'lantirish- urug'lanish, bunda bir xil organizm tomonidan hosil qilingan gametalar (tasmalar) birlashadi.
Embrion davri
Ajralish
Ajralish Bu zigotaning ketma-ket mitotik bo'linishlari seriyasidir, buning natijasida tuxum sitoplazmasining katta hajmi yadrolari bo'lgan ko'p sonli kichik hujayralarga bo'linadi. Parchalanish natijasida blastomerlar deb ataladigan hujayralar hosil bo'ladi. Yirilishning oddiy bo'linishdan farqi shundaki, yangi hosil bo'lgan blastomerlar hajmi kattalashmaydi. Bu interfazaning presintetik davrining yo'qolishi tufayli mumkin bo'ladi. Bunda interfazaning sintetik davri oldingi mitozning telofazasidan boshlanadi. Shunday qilib, blastomerlar soni asta-sekin o'sib boradi, lekin ularning umumiy hajmi deyarli o'zgarmaydi. Parchalanish paytida hujayralar sitoplazmasi hujayra membranasining invaginatsiyasining paydo bo'lishi bilan bo'linadi ( jo'yaklarni maydalash).
Amfibiya tuxumlarining bo'linishi (qurbaqa): 1 - ikki hujayrali bosqich; 2 - to'rt hujayrali bosqich; 3 - sakkiz hujayrali bosqich;
4 - sakkizdan o'n olti hujayrali bosqichga o'tish (hayvon qutbining hujayralari allaqachon bo'lingan va vegetativ qutb hujayralari
faqat parchalana boshlaydi); 5 - maydalashning keyingi bosqichi; 6 - blastula; 7 - kesmadagi blastula.
Maydalash jarayonining biologik ahamiyati: takror ishlab chiqarish tsikllari tufayli zigotaning genotipi ko'payadi; hujayra massasi keyingi o'zgarishlar uchun to'planadi; embrion bir hujayralidan ko'p hujayraliga aylanadi.
Blastomerlarning bo'linishi sodir bo'ladi sinxron Va sinxron bo'lmagan. Ko'pgina turlarda rivojlanishning boshidanoq asinxron bo'ladi, boshqalarida esa birinchi bo'linishdan keyin shunday bo'ladi.
Parchalanish xarakteri, birinchi navbatda, tuxumning tuzilishi, asosan, sarig'ining miqdori va sitoplazmada tarqalish xususiyatlari bilan belgilanadi. Shu munosabat bilan, maydalash usuliga ko'ra, tuxumning ikkita asosiy turi ajralib turadi: butunlay ezilgan va qisman ezilgan. Tuxumning sitoplazmasi blastomerlarga to'liq bo'linganda to'liq bo'linish deyiladi. Bo'lishi mumkin forma- barcha hosil bo'lgan blastomerlar bir xil o'lcham va shaklga ega (alitsital va izolesital tuxumlar uchun odatiy) va notekis— teng boʻlmagan oʻlchamdagi blastomerlar hosil boʻladi (sarigʻi oʻrtacha boʻlgan telolesital tuxumlarga xos). Kichik blastomerlar hayvon qutbida, yiriklari - embrionning vegetativ qutbi hududida paydo bo'ladi.

A - to'liq; B - qisman; B - diskoidal.
Qisman maydalash- tuxum sitoplazmasi blastomerlarga to'liq bo'linmagan bo'linish turi. Qisman boʻlinishning bir turi diskoidal boʻlib, unda yadro joylashgan hayvon qutbidagi sitoplazmaning sariqsiz qismigina parchalanadi. Sitoplazmaning parchalanishga uchragan maydoni deyiladi germinal disk. Ushbu turdagi maydalash ko'p miqdorda sarig'i (sudraluvchilar, qushlar, baliqlar) bo'lgan kuchli telolesital tuxumlarga xosdir.
Hayvonlarning turli guruhlari vakillarida parchalanish o'ziga xos xususiyatlarga ega, ammo u tuzilishga o'xshash tuzilish - blastulaning shakllanishi bilan yakunlanadi.
Blastula
Blastula- bir qavatli embrion. U hujayralar qatlami - blastoderma, bo'shliqni cheklovchi - blastokoldan iborat. Blastulalar blastomerlarning bir-biridan ajralib turishi tufayli parchalanishning dastlabki bosqichida shakllana boshlaydi. Olingan bo'shliq suyuqlik bilan to'ldiriladi. Blastulaning tuzilishi ko'p jihatdan bo'linish turiga bog'liq.
Koeloblastula(tipik blastula) bir xil parchalanish natijasida hosil bo'ladi. Bu katta blastokol (lancelet) bilan bir qatlamli pufakchaga o'xshaydi.
Amfiblastula telolesital tuxumlarni maydalash natijasida hosil bo'ladi; blastoderma turli oʻlchamdagi blastomeralardan qurilgan: hayvonlar qutbidagi mikromerlar va vegetativ qutbdagi makromerlar. Bunda blastokel hayvonlar qutbiga (amfibiyalar) qarab siljiydi.

1 - koeloblastula; 2 - amfiblastula; 3 - diskoblastula; 4 - blastokist; 5 - embrioblast; 6 - trofoblast.
Diskoblastula diskoidal maydalash natijasida hosil bo'ladi. Blastulaning bo'shlig'i germinal disk (qush) ostida joylashgan tor yoriqga o'xshaydi.
Blastotsist suyuqlik bilan to'ldirilgan bir qavatli pufakcha bo'lib, unda embrion (embrion rivojlanadi) va embrionni (sut emizuvchilar) oziqlanishini ta'minlaydigan trofoblast mavjud.

1 - ektoderma; 2 - endoderma; 3 - blastopor; 4 - gastrokoel.
Blastulaning paydo bo'lishidan keyin embriogenezning keyingi bosqichi boshlanadi - gastrulyatsiya(germ qatlamlarining shakllanishi). Gastrulyatsiya natijasida ikki qavatli, keyin esa uch qavatli embrion (ko'pchilik hayvonlarda) - gastrula hosil bo'ladi. Dastlab tashqi (ektoderma) va ichki (endoderma) qatlamlar hosil bo'ladi. Keyinchalik ekto- va endoderma o'rtasida uchinchi germ qatlami - mezoderma hosil bo'ladi.
Mikrob qatlamlari- embrionda ma'lum bir pozitsiyani egallagan va tegishli organlar va organ tizimlarini keltirib chiqaradigan hujayralarning alohida qatlamlari. Jinsiy qatlamlar nafaqat hujayra massalarining harakati natijasida, balki o'xshash, nisbatan bir hil blastula hujayralarining differensiallanishi natijasida ham paydo bo'ladi. Gastrulyatsiya jarayonida mikrob qatlamlari kattalar organizmining strukturaviy rejasiga mos keladigan pozitsiyani egallaydi. Differentsiatsiya- embrionning alohida hujayralari va qismlari o'rtasidagi morfologik va funktsional farqlarning paydo bo'lishi va ortishi jarayoni. Blastulaning turiga va hujayra harakatining xususiyatlariga qarab gastrulyatsiyaning quyidagi asosiy usullari farqlanadi: invaginatsiya, immigratsiya, delaminatsiya, epiboliya.

1 - invaginatsiya; 2 - epibolik; 3 - immigratsiya; 4 - delaminatsiya;
a - ektoderma; b - endoderma; c - gastrokoel.
Da invajinatsiya blastodermaning bo'limlaridan biri blastokelga (lansletda) kirib kela boshlaydi. Bunday holda, blastokel deyarli butunlay siljiydi. Ikki qavatli xalta hosil bo'lib, uning tashqi devori birlamchi ektoderma, ichki devori esa birlamchi endoderma bo'lib, birlamchi ichak bo'shlig'ini qoplaydi yoki gastrosel. Bo'shliq atrof-muhit bilan aloqa qiladigan teshik deyiladi blastopor, yoki asosiy og'iz. Hayvonlarning turli guruhlari vakillarida blastoporning taqdiri boshqacha. Protostomalarda u og'iz teshigiga aylanadi. Deyterostomalarda blastopor o'sib chiqadi va uning o'rnida tez-tez anal teshik paydo bo'ladi va og'iz teshiklari qarama-qarshi qutbdan (tananing oldingi uchi) yoriladi.
Immigratsiya- blastoderma hujayralarining bir qismini blastokel (yuqori umurtqalilar) bo'shlig'iga "quvib chiqarish". Bu hujayralardan endoderma hosil bo'ladi.
Delaminatsiya blastokolsiz blastulaga ega hayvonlarda (qushlarda) uchraydi. Ushbu gastrulyatsiya usuli bilan hujayra harakati minimal yoki umuman yo'q, chunki tabaqalanish sodir bo'ladi - blastulaning tashqi hujayralari ektodermaga aylanadi, ichki qismi esa endodermani hosil qiladi.
Epiboliya hayvonlar qutbining kichikroq blastomeralari tezroq parchalanib, o'simlik qutbning kattaroq blastomeralarini ko'paytirib, ektoderma (amfibiyalar) hosil qilganda yuzaga keladi. Vegetativ qutbning hujayralari ichki germ qatlamini - endodermani hosil qiladi.
Ta'riflangan gastrulyatsiya usullari kamdan-kam hollarda sof shaklda topiladi va ularning kombinatsiyasi odatda kuzatiladi (amfibiyalarda epiboliya bilan invaginatsiya yoki echinodermalarda immigratsiya bilan delaminatsiya).
Ko'pincha mezodermaning uyali moddasi endodermaning bir qismidir. U cho'ntak shaklidagi o'simtalar ko'rinishida blastokelga kirib boradi, so'ngra ular bog'lanadi. Mezoderma hosil bo'lganda, ikkinchi darajali tana bo'shlig'i yoki koelom hosil bo'ladi.
Embrion rivojlanishda organ hosil bo'lish jarayoni deyiladi organogenez. Organogenezni ikki bosqichga bo'lish mumkin: nevrulyatsiya- deyarli butun embrionni o'z ichiga olgan eksenel organlar majmuasi (asab naychasi, notokord, ichak naychasi va somit mezoderma) shakllanishi va boshqa organlarning qurilishi, tananing turli qismlari tomonidan ularning tipik shakli va ichki tashkiliy xususiyatlarini olish, ma'lum nisbatlarni o'rnatish (fazoda cheklangan jarayonlar).
tomonidan Karl Baerning mikrob qatlami nazariyasi, organlarning paydo bo'lishi u yoki bu mikrob qatlami - ekto-, mezo- yoki endodermaning o'zgarishi bilan bog'liq. Ayrim organlar aralash kelib chiqishi mumkin, ya'ni ular bir vaqtning o'zida bir nechta germ qatlamlari ishtirokida hosil bo'ladi. Masalan, ovqat hazm qilish traktining mushaklari mezodermaning hosilasi, ichki qoplamasi esa endoderma hosilasidir. Biroq, biroz soddalashtirish uchun asosiy organlar va ularning tizimlarining kelib chiqishi hali ham ma'lum mikrob qatlamlari bilan bog'liq bo'lishi mumkin. Neurulyatsiya bosqichidagi embrion deyiladi nevrula. Umurtqali hayvonlarning asab tizimini qurish uchun ishlatiladigan material - neyroektoderma, ektodermaning dorsal qismiga kiradi. U notokord rudimentining tepasida joylashgan.

1 - ekto-dermis; 2 - akkord; 3 - ikkilamchi tana bo'shlig'i; 4 - mezo-dermis; 5 - endo-dermis; 6 - ichak bo'shlig'i; 7 - asab naychasi.
Birinchidan, neyroektoderma hududida hujayra qatlamining tekislanishi sodir bo'ladi, bu esa asab plastinkasining shakllanishiga olib keladi. Keyin nerv plastinkasining qirralari qalinlashadi va ko'tariladi va asab burmalarini hosil qiladi. Plitaning markazida hujayralarning o'rta chiziq bo'ylab harakatlanishi tufayli embrionni kelajakdagi o'ng va chap yarmiga bo'linadigan neyron truba paydo bo'ladi. Nerv plastinkasi o'rta chiziq bo'ylab katlana boshlaydi. Uning qirralari tegib, keyin yopiladi. Ushbu jarayonlar natijasida bo'shliqli asab naychasi paydo bo'ladi - neyrokoelom.
Tizmalarning yopilishi birinchi navbatda o'rtada, so'ngra nerv yivining orqa qismida sodir bo'ladi. Nihoyat, bu boshqalardan ko'ra kengroq bo'lgan bosh qismida sodir bo'ladi. Oldingi kengaytirilgan qism qo'shimcha ravishda miyani hosil qiladi, asab naychasining qolgan qismi orqa miya hosil qiladi. Natijada, nerv plastinkasi ektoderma ostida yotgan nerv naychasiga aylanadi.
Neyrulyatsiya paytida nerv plastinkasining ba'zi hujayralari asab naychasining bir qismi emas. Ular nerv naychasi bo'ylab hujayralar to'plamini yoki ganglion plitasini yoki nerv tepasini hosil qiladi. Keyinchalik bu hujayralar embrion bo'ylab ko'chib o'tadi, asab ganglionlari, buyrak usti medullasi, pigment hujayralari va boshqalarni hosil qiladi.
Ektoderma materialidan nerv naychasidan tashqari epidermis va uning hosilalari (tuklar, sochlar, tirnoqlar, tirnoqlar, teri bezlari va boshqalar), ko'rish, eshitish, hidlash organlarining tarkibiy qismlari, og'iz epiteliysi va tish emalining rivojlanishi.
Mezodermal va endodermal organlar nerv naychasi hosil bo'lgandan keyin emas, balki u bilan bir vaqtda hosil bo'ladi. Birlamchi ichakning yon devorlari bo'ylab endodermaning chiqib ketishidan cho'ntaklar yoki burmalar hosil bo'ladi. Bu burmalar orasida joylashgan endoderma maydoni qalinlashadi, egiladi, burmalanadi va endodermaning asosiy massasidan ajralib chiqadi. Bu shunday ko'rinadi akkord. Hosil boʻlgan endodermaning choʻntaksimon oʻsimtalari birlamchi ichakdan ajraladi va bir qator segmental joylashgan yopiq qoplarga aylanadi, ularni selomik qoplar deb ham ataladi. Ularning devorlari mezoderma tomonidan hosil qilingan va ichidagi bo'shliq ikkilamchi tana bo'shlig'idir (yoki umuman).
Mezodermadan barcha turdagi biriktiruvchi to‘qima, teri, skelet, yo‘l-yo‘l va silliq muskullar, qon aylanish va limfa tizimlari, jinsiy sistema rivojlanadi.
Endodermadan ichak va oshqozon epiteliysi, jigar hujayralari, oshqozon osti bezining sekretsiya hujayralari, ichak va oshqozon bezlari rivojlanadi. Embrion ichakning oldingi bo'limi o'pka va nafas yo'llarining epiteliysini hosil qiladi, gipofizning oldingi va o'rta bo'laklari, qalqonsimon va paratiroid bezlarini sekretsiya qiladi.

1 - xordo-mezo-dermisning rudimenti; 2 - blastulaning bo'shlig'i; 3 — induktsiyalangan nerv naychasi; 4 - induksiyalangan akkord; 5 - asosiy nerv naychasi; 6 — asosiy akkord; 7 - mezbon embrion bilan bog'langan ikkilamchi embrionning shakllanishi.
- bu embrionning qismlari o'rtasidagi o'zaro ta'sir bo'lib, uning bir qismi - induktor - boshqa qism bilan aloqa qilish - reaksiyaga kirishuvchi tizim - ikkinchisining rivojlanish yo'nalishini belgilaydi.
Induksiya hodisasini 1901-yilda H.Spemann amfibiya embrionlarida ektodermal epiteliydan koʻz linzalari hosil boʻlishini oʻrganayotganda kashf etgan. 1924 yilda H. Spemann va G. Mangoldning tajribalari natijalari e'lon qilindi, ular embrion induksiya mavjudligining klassik isboti hisoblanadi. Erta gastrula bosqichida normal sharoitda asab tizimining tuzilmalariga aylanishi kerak bo'lgan ektodermaning rudimenti qorin bo'shlig'i ektodermasi ostiga tepalikli (pigmentsiz) triton embrionidan ko'chirildi, terining epidermisini, oddiy (pigmentli) triton embrionini keltirib chiqaradi. Natijada, nerv naychasi va eksenel organlar majmuasining boshqa tarkibiy qismlari dastlab qabul qiluvchi embrionning qorin tomonida paydo bo'ldi, keyin esa qo'shimcha embrion hosil bo'ldi. Bundan tashqari, kuzatishlar shuni ko'rsatdiki, qo'shimcha embrionning to'qimalari deyarli faqat qabul qiluvchining uyali materialidan hosil bo'ladi.
Agar notokord rudimenti gastrulaning dastlabki bosqichida butunlay olib tashlansa, asab naychasi rivojlanmaydi. Embrionning dorsal tomonidagi ektoderma, undan odatda nerv naychasi hosil bo'lib, teri epiteliysini hosil qiladi. Embrionlarning rivojlanishini keyingi o'rganish natijasida ma'lum bo'ldiki, xordomezoderma rudimenti asab naychasining induktori bo'lib, differentsiatsiya uchun asab tizimi rudimentidan induktor ta'sirini talab qiladi.
Postembrional rivojlanish davri
Rivojlanishning postembrional davri organizmning tug'ilish yoki tuxum membranalaridan chiqishi paytidan boshlanadi va uning o'limiga qadar davom etadi. Postembrional rivojlanish quyidagilarni o'z ichiga oladi: organizmning o'sishi; tananing yakuniy nisbatlarini belgilash; organ tizimlarining kattalar organizmi rejimiga o'tishi (xususan, balog'at yoshi). Farqlash Postembrional rivojlanishning ikkita asosiy turi: 1) to‘g‘ridan-to‘g‘ri, 2) transformatsiya bilan.
Da bevosita rivojlanish Individ onaning tanasidan yoki tuxum qobig'idan chiqadi, kattalar organizmidan faqat kichikroq hajmda (qushlar, sutemizuvchilar) farqlanadi.

Lar bor: lichinka bo'lmagan(tuxumdon) turi, bunda embrion tuxum (baliq, qush) ichida rivojlanadi; intrauterin embrion onaning tanasi ichida rivojlanib, platsenta (platsenta sut emizuvchilari) orqali u bilan bog'langan tur.
Transformatsiya (metamorfoz) bilan rivojlanish jarayonida tuxumdan lichinka paydo bo'ladi, bu kattalar hayvoniga qaraganda tuzilishi jihatidan oddiyroq (ba'zan undan juda farq qiladi); qoida tariqasida, u maxsus lichinka organlariga ega va ko'pincha kattalar hayvoniga (hasharotlar, ba'zi araxnidlar, amfibiyalar) qaraganda boshqacha turmush tarzini olib boradi.
Masalan, dumsiz amfibiyalarda tuxum qobig'idan lichinka, chuvalchang chiqadi. Tana shakli tekislangan, quyruq suzgichi, gill tirqishlari va gillalari, lateral chiziqli organlari, ikki kamerali yurak va bitta qon aylanishi mavjud. Vaqt o'tishi bilan, qalqonsimon gormon ta'siri ostida, tadpole metamorfozga uchraydi. Uning dumi eriydi, oyoq-qo'llari paydo bo'ladi, lateral chiziq yo'qoladi, o'pka va qon aylanishining ikkinchi doirasi rivojlanadi, ya'ni. asta-sekin u amfibiyalarga xos xususiyatlarni oladi.
Partenogenez
Partenogenez urug'lanmagan tuxumdan organizmning rivojlanishi deyiladi. U bir qator oʻsimlik turlarida, umurtqasizlar va umurtqali hayvonlarda uchraydi, sutemizuvchilardan tashqari, embrionogenezning dastlabki bosqichida partenogen embrionlar nobud boʻladi. Partenogenez sun'iy yoki tabiiy bo'lishi mumkin.
Sun'iy partenogenez odamning tuxumni turli moddalarga ta'sir qilish orqali faollashishi, mexanik tirnash xususiyati, haroratning oshishi va h.k.
Da tabiiy partenogenez tuxum faqat ichki yoki tashqi sabablar ta'sirida sperma ishtirokisiz parchalanib, embrionga aylana boshlaydi. Farqlash somatik Va generativ partenogenez. Generativ yoki haploid partenogenez davrida embrion gaploid tuxumdan (dron asalarilar) rivojlana boshlaydi. Somatik yoki diploid partenogenezda embrion diploid hujayradan rivojlana boshlaydi: 1) yoki diploid oositdan (meyoz bo'lmaydi), 2) yoki ikkita gaploid yadroning birlashishi natijasida hosil bo'lgan hujayradan (meyoz). uchraydi) (shira, dafniya, karahindiba).
Agar tuxumning rivojlanishi sperma yadrosi ishtirokisiz sodir bo'lsa (ba'zi baliqlar, yumaloq qurtlar), unda partenogenezning bu turi deyiladi. ginogenez. Biroq, bu tuxumni urug'lantirmasa ham, parchalanish boshlanishini rag'batlantiradigan sperma.
Agar tuxumning rivojlanishi faqat spermatozoidning genetik materiali va tuxum sitoplazmasi tufayli yuzaga kelsa, unda bu holda ular haqida gapirishadi. androgenez. Rivojlanishning bunday turi tuxumning yadrosi urug'lantirilgunga qadar o'lib qolsa va tuxumga bir emas, balki bir nechta sperma (ipak qurti) kirsa sodir bo'lishi mumkin.
ga boring 15-sonli ma'ruzalar"Angiospermlarda jinsiy ko'payish"
ga boring 17-sonli ma'ruzalar“Genetikaning asosiy tushunchalari. Mendel qonunlari"
Ko'p hujayrali organizmlarning ontogenezini davriylashtirish
Hayvonlarda germinal (embrion) bosqich va uning davrlari.
4. Embrional bosqichonaning tanasida yoki tuxum ichida yangi organizm paydo bo'ladigan vaqt. Embriogenez tug'ilish bilan tugaydi (tug'ilish, unib chiqish). Embrion davri urug'lantirilgandan yoki partenogenez davrida tuxum faollashganidan keyin boshlanadi va onaning tanasi, tuxumi, urug'i ichida sodir bo'ladi. Embrion rivojlanishi tug'ilish (sutemizuvchilar), tuxum qobig'idan (qushlar, sudraluvchilar) va unib chiqish (urug'li o'simliklar) bilan tugaydi. Embrion davrining asosiy bosqichlari bo'linish, gastrulyatsiya, gistogenez va organogenezdir.
Ajralish- zigotaning ketma-ket mitotik bo'linishlari ketma-ketligi, bu bir qavatli bosqich - blastulaning shakllanishi bilan yakunlanadi. Mitoz natijasida hujayralar soni ko'payadi, lekin interfaza juda qisqa va blastomerlar o'smaydi. Turli organizmlar guruhlarida maydalashning xususiyatlari sarig'ining joylashishi va miqdorining tabiatiga bog'liq bo'lib, shu munosabat bilan maydalashning ikki turi ajratiladi.
Gastrulyatsiya - Bu ikki qavatli embrion - gastrula hosil bo'lish jarayonidir. Gastrulyatsiya paytida hujayra o'sishi sodir bo'lmaydi. Bu bosqichda embrion tanasining ikki yoki uch qatlami - germ qatlamlari hosil bo'ladi. Gastrulyatsiya jarayonida ikki bosqichni farqlash nihoyatda muhimdir: a) ekto- va endodermaning shakllanishi (erta gastrula - ikki qavatli embrion) b) mezodermaning shakllanishi (kech gastrula hosil bo'ladi). - uch qavatli embrion). Gastrulyatsiya bosqichida ikki qavatli hayvonlar (gubkalar, koelenteratlar) embriogenezi tugallanadi, uch qavatli hayvonlarning embrion rivojlanishida mezoderma yotqiziladi (tekis chuvalchanglardan boshlab).
Turli organizmlarda gastrula turlicha shakllanadi. Gastrula shakllanishining quyidagi turlari ajratiladi: invaginatsiya (invaginatsiya), delaminatsiya (stratifikatsiya), epiboliya (ifloslanish), immigratsiya (o'rmalab ketish).
Gistogenez va organogenez - to'qimalar va organlarning shakllanishi. Bu jarayonlar differentsiatsiya (hujayralar, to'qimalar, organlarning tuzilishi va funktsiyalarida farqlarning paydo bo'lishi) tufayli amalga oshiriladi. O'quv to'qimalarining boshlang'ich hujayralari o'simliklar gistogenezida, hayvonlarning gistogenezida poya, yarim poya va etuk hujayralar ishtirok etadi. Organogenezda hujayralararo o'zaro ta'sirlar va biologik faol moddalarning ta'siri katta rol o'ynaydi. Gistogenez va organogenez fazalari (lancelet misolida) nevrulyatsiya - organlarning eksenel kompleksining shakllanishi (asab naychasi, notokord), boshqa organlarning shakllanishi - organlar kattalarga xos bo'lgan tizimli xususiyatlarga ega bo'ladi. Organogenez asosan rivojlanishning embrion davrining oxirida tugaydi, lekin postembriogenezda organlarning differentsiatsiyasi va asorati davom etadi.
Ko'p hujayrali organizmlar ontogenezini davriylashtirish - tushunchasi va turlari. "Ko'p hujayrali organizmlar ontogenezining davriyligi" toifasining tasnifi va xususiyatlari 2017, 2018 yil.

")







- Qishloq xo'jaligini rivojlantirish tendentsiyalari
- Yakovlev Aleksandr Sergeevich Aviatsiya sohasidagi ijodkorlikni baholash va Yakovlev bilan
- Knyaz Izyaslav I Izyaslav qaysi shaharda hukmronlik qilgan?
- Uyda olma sharbati - sharbat chiqargichsiz oddiy retsept
- Tuxumni sekin pishirgichda, bug'da, suvda va omlet sifatida qanday qaynatish mumkin?
- Sho'r suvda mazali tez pishiriladigan engil tuzlangan bodring
- Engil tuzlangan bodringni turli yo'llar bilan qanday tuzlash mumkin
- Sodani pishirish uchun sirka bilan qanday o'chirish kerak, qanday sirka kerak, uni nima bilan almashtirish mumkin
- Uy qurgan tovuqdan tovuq bulyoni
- Qanday qilib istalmagan odamni uydan uzoqroq tutish kerak?
- Okroshka - ovqat va idishlarni tayyorlash
- Oksidlarning fizik xossalari
- Cheat varaq: organizmlarning individual rivojlanishi (ontogenez)
- Nekrasov Nikolay Alekseevich Reja tuzing Nikolay Alekseevich Nekrasov
- Eng oddiy trigonometrik tenglamalar Trigonometrik tenglamalarni yechish usullari
- Trigonometrik tenglamalarni yechish Yordamchi burchak bilan tanishtirish
- Moddalar sinflari o'rtasidagi genetik bog'liqlik Ikki genetik seriya vakillari o'rtasidagi reaktsiyalar
- Yu. nomidagi Poltava milliy texnika universiteti
- Oy tutilishi haqida qiziqarli ma'lumotlar Quyoshni oyning qaysi fazasida kuzatish mumkin?
- "Qanday qilib" so'zidan oldin vergul bormi yoki yo'qmi?