برگه تقلب: رشد فردی ارگانیسم ها (آنتوژنز). مفهوم آنتوژنز
آنتوژنز(از یونانی όntos - موجود) یا رشد فردی -رشد یک فرد از لحظه تشکیل یک زیگوت یا جنین دیگر تا تکمیل طبیعی چرخه زندگی آن (تا زمان مرگ یا توقف وجود در ظرفیت قبلی). از نقطه نظر ژنتیکی، انتوژنز فرآیند آشکارسازی و اجرای اطلاعات ارثی جاسازی شده در سلول های زایای است.
Ontogenesis یک ویژگی جدایی ناپذیر از هر فرد است، مستقل از وابستگی سیستماتیک آن. بدون ظهور آنتوژن، تکامل حیات غیرقابل تصور خواهد بود. رشد فردی موجودات ارتباط نزدیکی با رشد تاریخی دارد - فیلوژنی(از یونانی phyle - قبیله).
انتوژنز افراد گونه های مختلف از نظر مدت زمان، سرعت و ماهیت تمایز متفاوت است. در حیوانات چند سلولی و انسان، شروع انتوژنز قبل از یک دوره است پیش جنینی رشد (پیش جنینی) - پیش زایی . در این دوره، سلول های زاینده تشکیل می شوند، فرآیند لقاح و تشکیل زیگوت رخ می دهد.
چهار دوره در انتوژنز وجود دارد: پیش جنینی، جنینی (قبل از تولد ), پس از جنینی (پس از زایمان ) و حالت بالغ از جمله پیری و مرگ در حیوانات، دوره جنینی معمولاً غنی از تمایز است و در گیاهان، دوره پس از جنین غنی است. هر یک از این دوره های انتوژنز به نوبه خود می توانند به مراحل کیفی متوالی تقسیم شوند.
پیش جنینیشامل گامتوژنز و لقاح است.
جنینیاین دوره با رشد جنین در محیط خارجی یا در دستگاه تناسلی بدن مادر و فرآیندهای سریع مورفوژنز مشخص می شود. در نتیجه این فرآیندها، یک ارگانیسم چند سلولی در مدت زمان کوتاهی ظاهر می شود.
در رشد جنین انسان سه دوره وجود دارد: ابتدایی , جنینی , جنینی (جنینی ).
ابتداییاین دوره هفته اول رشد جنینی را در بر می گیرد. از لحظه لقاح شروع می شود و تا لانه گزینی جنین در مخاط رحم ادامه می یابد.
جنینیدوره در انسان از لحظه لانه گزینی تا تکمیل فرآیند اندام زایی (2 تا 8 هفته) شروع می شود. این دوره با فرآیندهای اندام زایی، ویژگی های خاص تغذیه مشخص می شود - هیستیوتروفیک تغذیه، زمانی که جنین از ترشحات غدد رحمی و محصولات تجزیه بافت های مخاطی رحم تغذیه می کند. در این دوره رشد، گردش خون جفتی برای مدت طولانی وجود ندارد و ویژگی های مشخصه جنین انسان به دست می آید.
جنینییا دوره جنینی رشد جنین انسان از هفته نهم پس از لقاح آغاز می شود و تا زمان تولد ادامه می یابد. این دوره با افزایش رشد، فرآیندهای رشد سریع و ویژگی های تغذیه ای خاص مشخص می شود - هموتروفیک تغذیه ای که در ارتباط با عملکرد گردش خون جفت رخ می دهد. ویژگی های دوره های رشد جنین انسان در جدول 5 ارائه شده است .
جدول 5
ویژگی های دوره های رشد جنین انسان
پس از جنیندوره در انسان و پستانداران از لحظه تولد شروع می شود، از غشاهای جنینی خارج می شود تا پایان عمر و تا شروع بلوغ ادامه می یابد. در حیوانات تخمزا، این دوره از لحظه بیرون آمدن جوان از پوسته تخم شروع میشود. در گیاهان - از لحظه ای که ریشه اصلی ظاهر می شود.
انتقال به بدن بالغ می تواند به طور مستقیم یا غیر مستقیم انجام شود. در این راستا، سه نوع انتوژنز متمایز می شود: لارو , غیر لارو و داخل رحمی .
لارو، یا غیر مستقیم این نوع رشد مشخصه بسیاری از کولترها، کرم ها، نرم تنان، سخت پوستان، حشرات، نیزه ها، ریه ماهی و برخی از ماهیان استخوانی و دوزیستان است. این نوع رشد با وجود مراحل لاروی متمایز می شود. لاروها پس از بیرون آمدن از تخم، سبک زندگی فعالی را دنبال می کنند و خودشان غذا می گیرند. لاروها شبیه به شکل والدین نیستند - آنها از نظر ساختار بسیار ساده تر هستند، اندام های موقتی دارند که متعاقباً جذب می شوند (جذب می شوند) و در بزرگسالان مشاهده نمی شوند.
تحول بیشتر - دگردیسی - لاروها را می توان با توجه به نوع آنها به بالغ تبدیل کرد تحول کامل ، که در آن لارو به شدت با حشره بالغ تفاوت دارد و چندین مرحله رشد را طی می کند که مرحله اصلی آن مرحله شفیرگی (پروانه) است. یا رشد بدون مرحله شفیرگی - با توجه به نوع - رخ می دهد تحول ناقص و لارو خود شبیه یک حیوان بالغ است، اما از نظر اندازه کوچکتر است (ملخ، ملخ).
غیر لارو (سر راست ) نوع رشد با ظاهر ارگانیسمی شبیه به شکل والدین بزرگسال مشخص می شود، اما در اندازه کوچکتر و دستگاه تولید مثل کاملاً توسعه نیافته با آن متفاوت است. در چنین اشکالی از حیوانات (ماهی، خزندگان، پرندگان، پستانداران تخمزا، سرپایان، کلنترات) همه اندامها در طول دوره رشد جنینی تشکیل میشوند و رشد، بلوغ و تمایز عملکردها در دوره پس از جنینی اتفاق میافتد. رشد مستقیم با ذخیره زیاد زرده در تخم مرغ و وجود وسایل حفاظتی برای جنین در حال رشد یا با رشد جنین در بدن مادر همراه است.
داخل رحمی (سر راست ) جدیدترین نوع رشد از نظر فیلوژنتیک است. این مشخصه پستانداران عالی و انسان است که در آن تخم ها از نظر زرده ضعیف هستند و رشد جنین در رحم بدن مادر اتفاق می افتد. در این حالت اندام های موقت خارج جنینی تشکیل می شود که مهمترین آنها جفت است.
چرخه زندگی موجودات
چرخه زندگی، یا چرخه توسعه، شامل مراحل متوالی (اغلب مراحل نامیده می شود) است که مهمترین و کلیدی ترین حالات بدن را مشخص می کند - اصل و نسب , توسعه و تولید مثل .
در چرخه زندگی ارگانیسم های تولید مثل جنسی، دو مرحله وجود دارد: هاپلوئید و دیپلوئید . مدت نسبی این مراحل در بین نمایندگان گروه های مختلف موجودات زنده متفاوت است. بنابراین در تک یاخته ها و قارچ ها فاز هاپلوئید غالب است و در گیاهان و جانوران بالاتر فاز دیپلوئید غالب است.
طولانی شدن دیپلوفاز در طول تکامل با مزایای حالت دیپلوئید نسبت به حالت هاپلوئید توضیح داده می شود. به دلیل هتروزیگوسیتی و مغلوب بودن، آلل های مختلفی در حالت دیپلوئید حفظ و جمع می شوند. این مقدار اطلاعات ژنتیکی را در استخرهای ژنی جمعیتها و گونهها افزایش میدهد و منجر به تشکیل ذخیرهای از تنوع ارثی میشود که برای تکامل بیشتر امیدوارکننده است. در عین حال، در هتروزیگوت ها، آلل های مغلوب مضر بر رشد فنوتیپ تأثیر نمی گذارد و زنده ماندن موجودات را کاهش نمی دهد.
چرخه های زندگی وجود دارد ساده و مجتمع . پیچیدهها از چرخههای ساده تشکیل شدهاند، که در این مورد معلوم میشود که پیوندهای باز در یک چرخه پیچیده هستند.
تناوب نسلها مشخصه تقریباً همه جلبکهای پیشرفته تکاملی و همه گیاهان عالی است. یک نمودار کلی از چرخه زندگی یک گیاه که در آن تناوب نسل ها مشاهده می شود در شکل 1 ارائه شده است. یازده
برنج. 11. نمودار کلی چرخه زندگی گیاهی که دارای نسل های متناوب است
نمونه ای از گیاهان با چرخه ساده جلبک سبز تک سلولی کلرلا است که تنها توسط هاگ تکثیر می شود. رشد کلرلا با اتوسپورها شروع می شود. در حالی که هنوز داخل پوسته سلول مادر هستند، پوسته های خود را می پوشند و کاملاً شبیه به یک گیاه بالغ می شوند.
کلرلای جوان رشد می کند، به بلوغ می رسد و به عضوی از اسپورزایی تبدیل می شود - ظرف اختلاف نظر. در سلول مادر، 4-8 اتوسپور، دختر کلرلا، ظاهر می شود. در نتیجه، چرخه زندگی کلرلا به صورت دنباله ای از سه مرحله گرهی نشان داده می شود: موتوراسپرت → گیاه رویشی → سلول تولید مثل (ظرف) → موتوراسپرت و غیره.
بنابراین، یک چرخه زندگی ساده در طول تولیدمثل توسط هاگ دارای دنباله ای از سه مرحله گرهی است: 1 - یک پایه تک سلولی به عنوان مرحله اولیه گیاه، 2 - یک موجود تک سلولی یا چند سلولی بالغ، 3 - سلول مادر (تولید کننده) گیاه ابتدایی پس از مرحله سوم، سیر زندگی دوباره به مرحله پایه تک سلولی منتهی می شود.
چنین چرخه های زندگی ساده ای برای گیاهان معمولی نیست. اکثریت قریب به اتفاق گروه های گیاهی چرخه های زندگی پیچیده ای را نشان می دهند. آنها معمولاً شامل دو، گاهی اوقات سه چرخه ساده هستند. علاوه بر این، در چرخه های پیچیده (در طول تولید مثل جنسی) لزوماً 1-2 جداگانه وجود دارد فازهای گامت و زیگوت ها .
به عنوان مثال، یک سرخس همسپور در طبیعت با دو شکل از افراد نشان داده می شود - خود سرخس و رشد سرخس. پروتالوس سرخس (صفحات سبز کوچکی که به سختی روی خاک قابل مشاهده است) زاده مستقیم سرخس های بزرگ پنجه دار است. عمر کوتاهی دارد، اما می تواند زندگی یک فرد بزرگ برگ را ایجاد کند. در نتیجه، یک تغییر نسل وجود دارد: سرخس → پروتالوس → سرخس.
سرخس که توسط هاگ تولید مثل می کند نامیده می شود اسپوروفیت (نسل غیرجنسی)، و پروتالوس توسط گامت ها تولید مثل می کند و نامیده می شود گامتوفیت (نسل جنسی). گامتوفیت و اسپوروفیت فقط با روش تولید مثل فرد تعیین می شود. وجود جداگانه اسپوروفیت و گامتوفیت غیرممکن است و آنها فقط برای گیاهان با تناوب شدید نسل اعمال می شوند.
در آنژیوسپرم ها، گامتوفیت ماده معمولاً به هفت سلول کاهش می یابد، آرکگونیا ندارد و کیسه جنینی نامیده می شود. کیسه جنین، همولوگ با پروتالوس، از نظر میکروسکوپی کوچک است و در عمق گل قرار دارد.
گامتوفیت نر گیاهان دانه از یک میکروسپور ایجاد می شود و یک دانه گرده (گرده) است که به یک لوله گرده رشد می کند و دو سلول اسپرم را تشکیل می دهد. چرخه زندگی یک گیاه گلدار در شکل نشان داده شده است. 12.

برنج. 12. چرخه زندگی یک گیاه گلدار
اگر تولیدمثل جنسی با تولیدمثل پارتنوژنتیک و غیرجنسی جایگزین شود، چرخه های زندگی به طور قابل توجهی پیچیده تر می شوند. موجودات هاپلو دیپلوئیدی وجود دارند که در آنها یک جنس همیشه فقط در هاپلوفاز است و جنس دیگر در هر دو حالت دیپلو و هاپلوفاز. چنین موجوداتی شامل زنبور عسل است (شکل 13).

برنج. 13. چرخه زندگی زنبور عسل
سلول های سوماتیک رحم یک کلنی زنبور عسل دیپلوئید هستند و هاپلوفاز فقط توسط گامت ها نشان داده می شود. در یک زنبور کارگر، تخمدان ها کاهش یافته و هیچ هاپلوفازی در چرخه زندگی آن وجود ندارد. پهپادها به طور پارتنوژنتیکی از تخمهای بارور نشده رشد میکنند و دارای مجموعهای از کروموزومهای هاپلوئید هستند. به دلیل جایگزینی میوز توسط میتوز در گامتوژنز پهپادها، اسپرم آنها نیز هاپلوئید است. بنابراین، پهپادها فقط در هاپلوفاز وجود دارند.
قارچ ها به ویژه در چرخه زندگی خود متغیر هستند (شکل 14). در چرخه زندگی آنها، سه فاز هسته ای به وضوح تعریف شده است - هاپلوئید، دیپلوئید و دیکاریون.
دیکاریون در Ascomyces و Basidiomyces یافت می شود، در دومی بیشتر چرخه را تشکیل می دهد.
حالت هاپلوئید در بازیدیومیسس انتقالی است و حالت دیپلوئید فقط به صورت زیگوت وجود دارد.
در قارچ ها و جلبک ها نسبت طول مدت هاپلوفاز و دیپلوفاز تغییر می کند، بنابراین گونه های میانی مختلفی از چرخه های زندگی مشاهده می شود.

برنج. 14. طرح چرخه های زندگی اصلی قارچ ها
(تغییرات در فاز هسته ای با سایه های مختلف نشان داده می شود،
فلش ها جهت توسعه را نشان می دهند)
همبستگی فیلوژنی و انتوژنز 1. انتوژنز موجودات چند سلولی مسیر افزایش اندازه و پیچیدگی سازمان را دنبال می کند. 2. انتوژنز ارگانیسم بر اساس یک برنامه ژنتیکی مشخص به سمت هدف نهایی بلوغ و تولید مثل جنسی پیش می رود. 3. با گذار به چند سلولی، انتوژنز از نظر شکل پیچیده تر می شود و در زمان طولانی تر می شود. 4. ویژگی های کلی تکامل انتوژنز عبارتند از: n خودگردانی (برنامه ریزی) انتوژنز، n جهت تمایز آن، n از دست دادن دگردیسی، توالی تغییرات در برنامه های توسعه تحت تأثیر عوامل اپی ژنتیک. n
 رشد ریه ها در گروه های مختلف مهره داران: A - در axolotl (Ambystoma). ب - در اسپادفوت (Pelobates); ب - در وزغ (V/o)؛ G - مارمولک (Lacerta)، مراحل I-III رشد. سایه دوتایی قسمتهایی از ریه را نشان میدهد که تنها تحت تأثیر تنفس متمایز میشوند. مشاهده می شود که در وزغ و مارمولک تمایز قبل از شروع عملکرد رخ می دهد (طبق گفته A. A. Mashkovtsev، 1936)
رشد ریه ها در گروه های مختلف مهره داران: A - در axolotl (Ambystoma). ب - در اسپادفوت (Pelobates); ب - در وزغ (V/o)؛ G - مارمولک (Lacerta)، مراحل I-III رشد. سایه دوتایی قسمتهایی از ریه را نشان میدهد که تنها تحت تأثیر تنفس متمایز میشوند. مشاهده می شود که در وزغ و مارمولک تمایز قبل از شروع عملکرد رخ می دهد (طبق گفته A. A. Mashkovtsev، 1936)
 5. سازگاری هایی که در مراحل مختلف انتوژنز ظاهر می شوند و بقا را افزایش می دهند با انتخاب در طول نسل ها حفظ می شوند و عناصر رشد تاریخی گروهی از موجودات خواهند بود.
5. سازگاری هایی که در مراحل مختلف انتوژنز ظاهر می شوند و بقا را افزایش می دهند با انتخاب در طول نسل ها حفظ می شوند و عناصر رشد تاریخی گروهی از موجودات خواهند بود.
 طرح عارضه انتوژن ارگانیسم های چند سلولی در فرآیند تکامل تک سلولی. کلونی های تک سلولی نوع Volvox [سلول ها به جنسی (سیاه) و جسمی تمایز می یابند]. یک ارگانیسم چند سلولی از نوع هیدرا (مراحل بلاستولا و گاسترولا اضافه می شود). حیوان متقارن دو طرفه اولیه (مزودرم اضافه شده است). بالاترین حیوان متقارن دو طرفه.
طرح عارضه انتوژن ارگانیسم های چند سلولی در فرآیند تکامل تک سلولی. کلونی های تک سلولی نوع Volvox [سلول ها به جنسی (سیاه) و جسمی تمایز می یابند]. یک ارگانیسم چند سلولی از نوع هیدرا (مراحل بلاستولا و گاسترولا اضافه می شود). حیوان متقارن دو طرفه اولیه (مزودرم اضافه شده است). بالاترین حیوان متقارن دو طرفه.
 قانون تشابه ژرمینال "در انتوژنز حیوانات ابتدا ویژگی های گروه های طبقه بندی بالاتر ظاهر می شود و در فرآیند رشد جنینی ویژگی های دسته بندی طبقه بندی پایین تر تشکیل می شود" (K. Baer) این به دلیل محافظه کاری آن قسمت از ژنوم است. که مسئول فرآیندهای مورفوژنتیک در انتوژنز است. اما در فرآیند فیلوژنز، در نتیجه جهش های جنینی، جریان ژن ها تازگی، ویژگی های جدیدی به دست می آورد، یعنی ژنوتیپ های فرزندان با ژنوتیپ های اجدادشان متفاوت است.
قانون تشابه ژرمینال "در انتوژنز حیوانات ابتدا ویژگی های گروه های طبقه بندی بالاتر ظاهر می شود و در فرآیند رشد جنینی ویژگی های دسته بندی طبقه بندی پایین تر تشکیل می شود" (K. Baer) این به دلیل محافظه کاری آن قسمت از ژنوم است. که مسئول فرآیندهای مورفوژنتیک در انتوژنز است. اما در فرآیند فیلوژنز، در نتیجه جهش های جنینی، جریان ژن ها تازگی، ویژگی های جدیدی به دست می آورد، یعنی ژنوتیپ های فرزندان با ژنوتیپ های اجدادشان متفاوت است.

 پدیده شباهت ژرمینال. جنین همه مهره داران در مراحل اولیه رشد بیشتر از مراحل بعدی به یکدیگر شباهت دارند (به گفته E. Haeckel)
پدیده شباهت ژرمینال. جنین همه مهره داران در مراحل اولیه رشد بیشتر از مراحل بعدی به یکدیگر شباهت دارند (به گفته E. Haeckel)
 وراثت و تنوع در تکامل انتوژنز خود را به شکل خلاصهسازی، پالینژنز، سنوژنز، هتروکرونی (شتاب، عقب ماندگی)، هتروتوپی و فیلمبریوژنز نشان میدهد.
وراثت و تنوع در تکامل انتوژنز خود را به شکل خلاصهسازی، پالینژنز، سنوژنز، هتروکرونی (شتاب، عقب ماندگی)، هتروتوپی و فیلمبریوژنز نشان میدهد.
 F. Muller معتقد بود که بازآرایی های تکاملی می تواند دو نوع باشد. 1 رویه زایی فرزندان می تواند فراتر از مرحله ای که در اجداد به پایان رسیده است ادامه یابد. 2 رویه زایی نوادگان ممکن است در مراحل اولیه یا میانی از مسیری که در میان اجداد خود طی کرده است منحرف شود. قانون بیوژنتیک انتوژنیک ارگانیسم عبارت است از تکرار کوتاه و فشرده (بازنویسی) فیلوژنی یک گونه معین (E. Haeckel). A. N. Severtsov - 1. در فرآیند انتوژنز، ویژگی های مراحل رشد اشکال اجدادی بالغ ممکن است تکرار نشود. 2. در فرآیند انتوژنز می توان مسیرهای فیلوژنتیک جدیدی ایجاد کرد. به تغییراتی که در روند انتوژنز اهمیت فیلوژنتیکی میدهند، فیلمبروژنز میگویند.
F. Muller معتقد بود که بازآرایی های تکاملی می تواند دو نوع باشد. 1 رویه زایی فرزندان می تواند فراتر از مرحله ای که در اجداد به پایان رسیده است ادامه یابد. 2 رویه زایی نوادگان ممکن است در مراحل اولیه یا میانی از مسیری که در میان اجداد خود طی کرده است منحرف شود. قانون بیوژنتیک انتوژنیک ارگانیسم عبارت است از تکرار کوتاه و فشرده (بازنویسی) فیلوژنی یک گونه معین (E. Haeckel). A. N. Severtsov - 1. در فرآیند انتوژنز، ویژگی های مراحل رشد اشکال اجدادی بالغ ممکن است تکرار نشود. 2. در فرآیند انتوژنز می توان مسیرهای فیلوژنتیک جدیدی ایجاد کرد. به تغییراتی که در روند انتوژنز اهمیت فیلوژنتیکی میدهند، فیلمبروژنز میگویند.
 1. آنابولیا - گسترش مراحل پایانی مورفوژنز، یعنی فرآیند مورفوژنز پس از رسیدن به مرحله ای که مورفوژنز در اجداد به پایان می رسد ادامه می یابد. 2. انحراف - یک انحراف، تغییر در مسیر توسعه در مراحل میانی مورفوژنز. 3. Archalaxis - تغییرات در مراحل اولیه تشکیل یک اندام، یعنی پایه های آن. مورفوژنز از همان ابتدا مانند اجداد پیش نمی رود.
1. آنابولیا - گسترش مراحل پایانی مورفوژنز، یعنی فرآیند مورفوژنز پس از رسیدن به مرحله ای که مورفوژنز در اجداد به پایان می رسد ادامه می یابد. 2. انحراف - یک انحراف، تغییر در مسیر توسعه در مراحل میانی مورفوژنز. 3. Archalaxis - تغییرات در مراحل اولیه تشکیل یک اندام، یعنی پایه های آن. مورفوژنز از همان ابتدا مانند اجداد پیش نمی رود.
 انحراف و آرالاکسیس رشد فلس های استخوانی و مو: الف - فلس های استخوانی ماهی. ب - فلس های شاخی خزندگان؛ ب - موی پستانداران. فلش های منفرد - آنابولیا، از A به B - انحراف، از B به C - ■ archallaxis. هنگامی که مو ظاهر می شود، گروهی از سلول های اپیدرمی اولیه بیرون نمی زند، بلکه به داخل پوست فرود می آید؛ متعاقباً، کل رشد پایه رشد فیلوژنتیکی پوسته ها را تکرار نمی کند (طبق گفته A. N. Severtsov، 1939)
انحراف و آرالاکسیس رشد فلس های استخوانی و مو: الف - فلس های استخوانی ماهی. ب - فلس های شاخی خزندگان؛ ب - موی پستانداران. فلش های منفرد - آنابولیا، از A به B - انحراف، از B به C - ■ archallaxis. هنگامی که مو ظاهر می شود، گروهی از سلول های اپیدرمی اولیه بیرون نمی زند، بلکه به داخل پوست فرود می آید؛ متعاقباً، کل رشد پایه رشد فیلوژنتیکی پوسته ها را تکرار نمی کند (طبق گفته A. N. Severtsov، 1939)
 ظهور، دگرگونی و ناپدید شدن اندام ها در فیلوژنز چند کارکردی اندام ها و توانایی آنها در تغییر کمی عملکرد آنها زمینه ساز دگرگونی فیلوژنتیکی اندام ها است. 1. هر چه یک اندام وظایف بیشتری انجام دهد، جهت های بیشتری می تواند در طول تکامل تغییر کند. 2. چند کارکردی امکان دگرگونی تطبیقی سیستم ها را فراهم می کند، 3. وجود یا عدم وجود عملکردها جهت بازسازی اندام را در هنگام تغییر محیط تعیین می کند. 4. توابع اولیه و ثانویه وجود دارد. 5. تغییرات کمی در توابع ناشی از ناهمگنی ژنتیکی و تفاوت در تعداد و اندازه واحدهای مورفوعملکردی همگن یک اندام (بافت) است.
ظهور، دگرگونی و ناپدید شدن اندام ها در فیلوژنز چند کارکردی اندام ها و توانایی آنها در تغییر کمی عملکرد آنها زمینه ساز دگرگونی فیلوژنتیکی اندام ها است. 1. هر چه یک اندام وظایف بیشتری انجام دهد، جهت های بیشتری می تواند در طول تکامل تغییر کند. 2. چند کارکردی امکان دگرگونی تطبیقی سیستم ها را فراهم می کند، 3. وجود یا عدم وجود عملکردها جهت بازسازی اندام را در هنگام تغییر محیط تعیین می کند. 4. توابع اولیه و ثانویه وجود دارد. 5. تغییرات کمی در توابع ناشی از ناهمگنی ژنتیکی و تفاوت در تعداد و اندازه واحدهای مورفوعملکردی همگن یک اندام (بافت) است.
 روش های اساسی تبدیل اندام ها و عملکرد آنها. 1. تقویت عملکرد اصلی هر اندام به دو صورت انجام می شود: الف) با تغییر ساختار اندام، ب) یا با افزایش تعداد اجزای عملکردی در اندام.
روش های اساسی تبدیل اندام ها و عملکرد آنها. 1. تقویت عملکرد اصلی هر اندام به دو صورت انجام می شود: الف) با تغییر ساختار اندام، ب) یا با افزایش تعداد اجزای عملکردی در اندام.
 2. تضعیف عملکرد اصلی. ارگانیسم یک طرح بهینه را از نظر ساختار و عملکرد نشان می دهد. این مصلحت در نتیجه کمبود مزمن منابع غذایی و رقابت برای بقا به وجود می آید. فیزیولوژیستهای آمریکایی، تیلور و وایبل، اصل سمورفوزیس را فرموله کردند؛ قدرت هیچ ساختار کاری بیش از سطح مورد نیاز بدن تحت حداکثر بار است. 3. کاهش تعداد توابع. اندام ها یا ساختارهای تخصصی برخی از عملکردهای خود را از دست می دهند. 4. افزایش تعداد توابع. آ. افزایش تعداد عملکردهای اندام های فردی امکان دگرگونی های مورفوفانشنال را افزایش می دهد، ب. هنگامی که تعداد توابع افزایش می یابد، اصلی، به عنوان یک قاعده، تغییر نمی کند، اما توسط دیگران تکمیل می شود.
2. تضعیف عملکرد اصلی. ارگانیسم یک طرح بهینه را از نظر ساختار و عملکرد نشان می دهد. این مصلحت در نتیجه کمبود مزمن منابع غذایی و رقابت برای بقا به وجود می آید. فیزیولوژیستهای آمریکایی، تیلور و وایبل، اصل سمورفوزیس را فرموله کردند؛ قدرت هیچ ساختار کاری بیش از سطح مورد نیاز بدن تحت حداکثر بار است. 3. کاهش تعداد توابع. اندام ها یا ساختارهای تخصصی برخی از عملکردهای خود را از دست می دهند. 4. افزایش تعداد توابع. آ. افزایش تعداد عملکردهای اندام های فردی امکان دگرگونی های مورفوفانشنال را افزایش می دهد، ب. هنگامی که تعداد توابع افزایش می یابد، اصلی، به عنوان یک قاعده، تغییر نمی کند، اما توسط دیگران تکمیل می شود.
 5. تغییر توابع معمولاً زمانی رخ می دهد که شرایط وجود تغییر کند. اصلی ممکن است اهمیت خود را از دست بدهد و یکی از کارکردهای ثانویه ممکن است اهمیت اصلی را به دست آورد. 6. پلیمریزاسیون اندام ها. 7. الیگومریزاسیون اندام ها و تمرکز توابع
5. تغییر توابع معمولاً زمانی رخ می دهد که شرایط وجود تغییر کند. اصلی ممکن است اهمیت خود را از دست بدهد و یکی از کارکردهای ثانویه ممکن است اهمیت اصلی را به دست آورد. 6. پلیمریزاسیون اندام ها. 7. الیگومریزاسیون اندام ها و تمرکز توابع

 پیچیدگی روابط تکاملی در جایگزینی، هتروباتمی و جبران عملکردها آشکار می شود. جایگزینی - جایگزینی اندام ها و عملکردها در فرآیند انتوژنز و فیلوژنز توسط اندام دیگری الف) جایگزینی هموتوپیک. ب) جایگزینی هتروتوپیک. در هتروباتمی، تکامل سیستمها و اندامهای فردی با سرعتهای متفاوتی اتفاق میافتد. هتروباتمی سرعت های مختلف تخصصی شدن تکامل موزاییک سیستم های اندام. جبران - تاخیر در تکامل برخی از اندام ها را می توان با تغییرات سریع در سایر اندام ها جبران کرد.
پیچیدگی روابط تکاملی در جایگزینی، هتروباتمی و جبران عملکردها آشکار می شود. جایگزینی - جایگزینی اندام ها و عملکردها در فرآیند انتوژنز و فیلوژنز توسط اندام دیگری الف) جایگزینی هموتوپیک. ب) جایگزینی هتروتوپیک. در هتروباتمی، تکامل سیستمها و اندامهای فردی با سرعتهای متفاوتی اتفاق میافتد. هتروباتمی سرعت های مختلف تخصصی شدن تکامل موزاییک سیستم های اندام. جبران - تاخیر در تکامل برخی از اندام ها را می توان با تغییرات سریع در سایر اندام ها جبران کرد.
 آتاویسم ها ظهور در موجودات منفرد از نوع خاصی از ویژگی ها هستند که در اجداد آنها وجود داشته است، اما در طول فرآیند تکامل از بین رفته است. ریشه ها اندام های توسعه نیافته ای هستند که در مقایسه با اندام های همولوگ اشکال اجدادی عملاً عملکرد خود را در فرآیند تکامل از دست داده اند. ابتدایی سازی اندام ها. کاهش تعداد عملکردها در طول تکامل می تواند منجر به ضعیف شدن رشد یک اندام شود. ابتدایی ها معمولاً عملکردی دارند که معمولاً ربطی به عملکرد اصلی اندام ندارد. ریشه ها اغلب یک پیوند فعال در فرآیندهای مورفوژنتیک هستند که تشکیل طبیعی سایر اندام ها را تعیین می کنند. ابتدایی شدن می تواند به دو صورت اتفاق بیفتد: 1. انتوژنز یک اندام به همان روشی که در اجدادش انجام می شود، ادامه می یابد، اما در برخی مراحل متوقف می شود. 2 درد اندام در انتوژنز کمتر از اجداد است یا دیرتر رخ می دهد در نتیجه زمانی برای رشد ندارد.
آتاویسم ها ظهور در موجودات منفرد از نوع خاصی از ویژگی ها هستند که در اجداد آنها وجود داشته است، اما در طول فرآیند تکامل از بین رفته است. ریشه ها اندام های توسعه نیافته ای هستند که در مقایسه با اندام های همولوگ اشکال اجدادی عملاً عملکرد خود را در فرآیند تکامل از دست داده اند. ابتدایی سازی اندام ها. کاهش تعداد عملکردها در طول تکامل می تواند منجر به ضعیف شدن رشد یک اندام شود. ابتدایی ها معمولاً عملکردی دارند که معمولاً ربطی به عملکرد اصلی اندام ندارد. ریشه ها اغلب یک پیوند فعال در فرآیندهای مورفوژنتیک هستند که تشکیل طبیعی سایر اندام ها را تعیین می کنند. ابتدایی شدن می تواند به دو صورت اتفاق بیفتد: 1. انتوژنز یک اندام به همان روشی که در اجدادش انجام می شود، ادامه می یابد، اما در برخی مراحل متوقف می شود. 2 درد اندام در انتوژنز کمتر از اجداد است یا دیرتر رخ می دهد در نتیجه زمانی برای رشد ندارد.
 نمونه هایی از اندام های وستیژیال: A- اندام های عقبی مار پیتون (Python regius). بال B کیوی (Apteryx australis); B-عناصر کمربند لگنی نهنگ راست (Eubalaena glacialis) (طبق گفته St. Skovron، 1965؛ A. A. Paramonov، 1978
نمونه هایی از اندام های وستیژیال: A- اندام های عقبی مار پیتون (Python regius). بال B کیوی (Apteryx australis); B-عناصر کمربند لگنی نهنگ راست (Eubalaena glacialis) (طبق گفته St. Skovron، 1965؛ A. A. Paramonov، 1978
 دگرگونی های همبستگی اندام ها. یکپارچگی و ثبات آنتوژنز در رشد فردی و تاریخی به شکل همبستگی و هماهنگی فرآیندهای ریخت زایی آشکار می شود. همبستگی ها روابط عملکردی و ساختاری بین بخش های یک ارگانیسم در حال رشد هستند. همبستگی های ژنومی بر اساس تعامل و پیوند ژن ها در یک ژنوتیپ. همبستگی های مورفوژنتیک بر اساس تعامل سلول ها یا قسمت هایی از بدن با یکدیگر در طول جنین زایی است. همبستگی های ارگونتیک وابستگی های عملکردی بین ساختارهای قطعی ایجاد می کند. هماهنگی. تغییرات مرتبط در اندام ها در فیلوژنز. هماهنگی توپوگرافی اتصالات فضایی اندامها. هماهنگی پویا تغییرات در روند فیلوژنز اندام های به هم پیوسته عملکردی و سیستم های آنها. هماهنگی بیولوژیکی تغییرات تکاملی در اندام هایی که مستقیماً به یکدیگر مرتبط نیستند. انتخاب به سمت تغییر هماهنگ آنها برای اطمینان از زندگی حرکت می کند
دگرگونی های همبستگی اندام ها. یکپارچگی و ثبات آنتوژنز در رشد فردی و تاریخی به شکل همبستگی و هماهنگی فرآیندهای ریخت زایی آشکار می شود. همبستگی ها روابط عملکردی و ساختاری بین بخش های یک ارگانیسم در حال رشد هستند. همبستگی های ژنومی بر اساس تعامل و پیوند ژن ها در یک ژنوتیپ. همبستگی های مورفوژنتیک بر اساس تعامل سلول ها یا قسمت هایی از بدن با یکدیگر در طول جنین زایی است. همبستگی های ارگونتیک وابستگی های عملکردی بین ساختارهای قطعی ایجاد می کند. هماهنگی. تغییرات مرتبط در اندام ها در فیلوژنز. هماهنگی توپوگرافی اتصالات فضایی اندامها. هماهنگی پویا تغییرات در روند فیلوژنز اندام های به هم پیوسته عملکردی و سیستم های آنها. هماهنگی بیولوژیکی تغییرات تکاملی در اندام هایی که مستقیماً به یکدیگر مرتبط نیستند. انتخاب به سمت تغییر هماهنگ آنها برای اطمینان از زندگی حرکت می کند
 ناهنجاری های مادرزادی نقص های رشدی ناهنجاری های ساختاری هستند که قبل از تولد رخ می دهند، بلافاصله تشخیص داده می شوند یا مدتی پس از تولد باعث اختلال در عملکرد اندام می شوند. I. ناهنجاری های مادرزادی بسته به علت به 1. ارثی (ناشی از تغییرات در ژن ها یا کروموزوم ها که باعث اختلال در فرآیندهای بیوشیمیایی، سلولی، بافت و اندام و ارگانیسم می شود) تقسیم می شوند. 2. اگزوژن (به وجود آمده تحت تأثیر عوامل محیطی تراتوژن). از آنجایی که تراتوژن ها بر فرآیندهای مشابه جهش ها تأثیر می گذارند، تظاهرات فنوتیپی نقص های برون زا و ژنتیکی می تواند بسیار مشابه باشد که با اصطلاح فنوکپی مشخص می شود. 3. نقایص چند عاملی ناشی از تأثیر عوامل برون زا و ژنتیکی است.
ناهنجاری های مادرزادی نقص های رشدی ناهنجاری های ساختاری هستند که قبل از تولد رخ می دهند، بلافاصله تشخیص داده می شوند یا مدتی پس از تولد باعث اختلال در عملکرد اندام می شوند. I. ناهنجاری های مادرزادی بسته به علت به 1. ارثی (ناشی از تغییرات در ژن ها یا کروموزوم ها که باعث اختلال در فرآیندهای بیوشیمیایی، سلولی، بافت و اندام و ارگانیسم می شود) تقسیم می شوند. 2. اگزوژن (به وجود آمده تحت تأثیر عوامل محیطی تراتوژن). از آنجایی که تراتوژن ها بر فرآیندهای مشابه جهش ها تأثیر می گذارند، تظاهرات فنوتیپی نقص های برون زا و ژنتیکی می تواند بسیار مشابه باشد که با اصطلاح فنوکپی مشخص می شود. 3. نقایص چند عاملی ناشی از تأثیر عوامل برون زا و ژنتیکی است.
 II. بسته به مرحله انتوژنز قبل از تولد: 1. گامتوپاتی (اختلالات رشدی در مرحله زیگوت) 2. بلاستوپاتی (اختلالات رشدی در مرحله بلاستولا) 3. جنین (اختلالات در دوره 15 روزه تا 8 هفته) 4. جنین (اختلال) که پس از 10 هفته رخ داده است) نقض مورفوژنز جنینی (3-10 هفته) اغلب منجر به نقص در نتیجه اختلالات می شود: تولید مثل (هیپوپلازی و آپلازی اندام ها)، مهاجرت (هتروتوپی)، تمایز (تداوم ساختارهای جنینی، آپلازی). یک عضو یا بخشی از آن)، چسبندگی و مرگ سلولی (دیسرافیسم غیر همجوشی).
II. بسته به مرحله انتوژنز قبل از تولد: 1. گامتوپاتی (اختلالات رشدی در مرحله زیگوت) 2. بلاستوپاتی (اختلالات رشدی در مرحله بلاستولا) 3. جنین (اختلالات در دوره 15 روزه تا 8 هفته) 4. جنین (اختلال) که پس از 10 هفته رخ داده است) نقض مورفوژنز جنینی (3-10 هفته) اغلب منجر به نقص در نتیجه اختلالات می شود: تولید مثل (هیپوپلازی و آپلازی اندام ها)، مهاجرت (هتروتوپی)، تمایز (تداوم ساختارهای جنینی، آپلازی). یک عضو یا بخشی از آن)، چسبندگی و مرگ سلولی (دیسرافیسم غیر همجوشی).

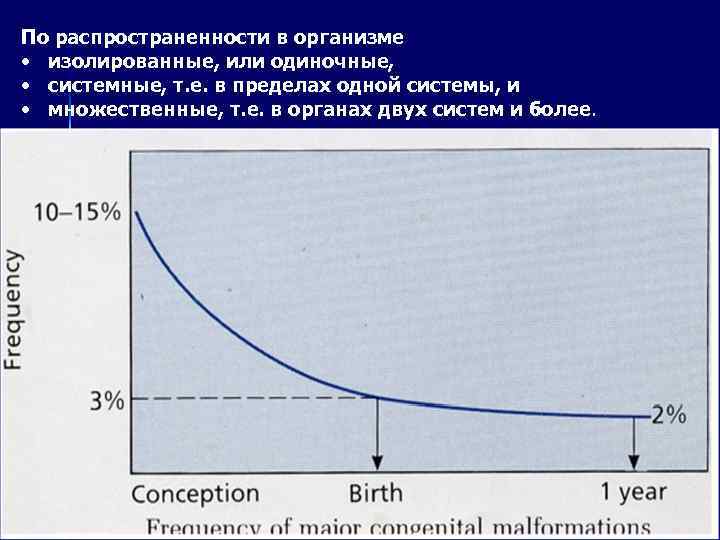 با توجه به شیوع آنها در بدن، منفرد یا منفرد، سیستمیک، یعنی در یک سیستم، و چندگانه، یعنی در اندام های دو یا چند سیستم.
با توجه به شیوع آنها در بدن، منفرد یا منفرد، سیستمیک، یعنی در یک سیستم، و چندگانه، یعنی در اندام های دو یا چند سیستم.
 اتیولوژی ناهنجاری های مادرزادی سیستمیک ایدئوپاتیک 60% مولتی فاکتوریال 20% مونوژنیک 7. 5% کروموزومی 6% بیماری های مادر 3% عفونت های داخل رحمی 2% داروها، اشعه، الکل و غیره 1. 5%
اتیولوژی ناهنجاری های مادرزادی سیستمیک ایدئوپاتیک 60% مولتی فاکتوریال 20% مونوژنیک 7. 5% کروموزومی 6% بیماری های مادر 3% عفونت های داخل رحمی 2% داروها، اشعه، الکل و غیره 1. 5%
 با توجه به اهمیت فیلوژنتیکی، ناهنجاری ها را می توان به دو دسته فیلوژنتیک و غیر فیلوژنتیک تقسیم کرد. نقصهای تعیینشده فیلوژنتیکی شبیه اندامهای حیوانات از شاخه Chordates و زیرگروه مهرهداران (نقص اجدادی یا آتاویستیک) است. آنها ارتباط ژنتیکی انسان را با سایر مهره داران نشان می دهند و همچنین به درک مکانیسم های وقوع نقص کمک می کنند. مکانیسم های اصلی برای وقوع آتاویسم احتمالاً جهش ژن های تنظیمی است که سرعت مورفوژنز و راه اندازی فرآیندهایی را با هدف کاهش اندام کنترل می کنند. رایجترین آتاویسمها عبارتند از: رشد ناکافی اندامها در مراحل مورفوژنز، زمانی که حالت اجدادی را جمعبندی کردهاند. تداوم ساختارهای جنینی، همچنین ویژگی مورفولوژی اجداد را خلاصه می کند. نقض حرکت اندام ها در انتوژنز.
با توجه به اهمیت فیلوژنتیکی، ناهنجاری ها را می توان به دو دسته فیلوژنتیک و غیر فیلوژنتیک تقسیم کرد. نقصهای تعیینشده فیلوژنتیکی شبیه اندامهای حیوانات از شاخه Chordates و زیرگروه مهرهداران (نقص اجدادی یا آتاویستیک) است. آنها ارتباط ژنتیکی انسان را با سایر مهره داران نشان می دهند و همچنین به درک مکانیسم های وقوع نقص کمک می کنند. مکانیسم های اصلی برای وقوع آتاویسم احتمالاً جهش ژن های تنظیمی است که سرعت مورفوژنز و راه اندازی فرآیندهایی را با هدف کاهش اندام کنترل می کنند. رایجترین آتاویسمها عبارتند از: رشد ناکافی اندامها در مراحل مورفوژنز، زمانی که حالت اجدادی را جمعبندی کردهاند. تداوم ساختارهای جنینی، همچنین ویژگی مورفولوژی اجداد را خلاصه می کند. نقض حرکت اندام ها در انتوژنز.




رویه زایی افراد گونه های مختلف از نظر مدت زمان، سرعت و ماهیت تمایز یکسان نیست (به زیر مراجعه کنید). معمولاً به دوره های پیش جنینی، جنینی و پس از جنین تقسیم می شود. در حیوانات، دوره جنینی معمولاً غنی از تمایز است، در گیاهان، دوره پس از جنین غنی است. هر یک از این دوره های انتوژنز را می توان به مراحل کیفی متوالی تقسیم کرد. آنتوژنز را می توان با توسعه مستقیم یا توسعه با دگردیسی مشخص کرد.
ویژگی های آنتوژنز در گروه های مختلف.اشکال تجلی فردیت در طبیعت زنده متنوع است و روند آنتوژنز در نمایندگان مختلف پروکاریوت ها، قارچ ها، گیاهان و حیوانات از نظر محتوا نابرابر است.
برنج. 14.1. طرح عارضه پی در پی انتوژن موجودات چند سلولی در فرآیند تکامل. الف - تولید مثل موجودات تک سلولی آزاد. ب - انتوژنز یک کلنی از نوع Volvox تک سلولی [سلول ها به جنسی (سیاه) و جسمی تمایز می یابند]. ب - انتوژنیک ارگانیسم چند سلولی از نوع هیدرا (مراحل بلاستولا و گاسترولا اضافه می شود). D - انتوژنز حیوان متقارن دو طرفه اولیه (مزودرم اضافه شده است). D - انتوژنز بالاترین حیوان متقارن دو طرفه (طبق گفته A.N. Severtsov، 1935)
با انتقال به چند سلولی (Metazoa)، انتوژنز از نظر شکل پیچیدهتر میشود و در زمان طولانیتر میشود (شکل 14.1)، اما در روند تکامل آنتوژنز، مواردی از سادهسازی توسعه نیز مشاهده میشود که با ظهور موارد بیشتری همراه است. روش های پیشرفته برای درک اطلاعات ارثی در طول تکامل، گیاهان و حیوانات چرخه های رشد پیچیده ای را تجربه می کنند که هر مرحله از آن با شرایط محیطی خاصی سازگار است. گاهی اوقات در فرآیند تکامل یک ساده سازی ثانویه چرخه های زندگی رخ می دهد.
با سادهسازی چرخه زندگی، کل فرآیند رشد انتوژنتیکی از نظر کیفی تغییر میکند. یکی از پیامدهای سادهسازی چرخه زندگی، گذار از مرحله رشد هاپلوئید به مرحله دیپلوئید و از رشد با دگردیسی (مثلاً در دوزیستان) به رشد مستقیم (در خزندگان و سایر مهرهداران برتر) است. با رشد مستقیم، یک حیوان تازه متولد شده تمام ویژگی های سازمانی اولیه یک موجود بالغ را دارد. توسعه با دگردیسی از طریق یک سری مراحل لاروی انجام می شود. یک لارو از تخم بیرون می آید و از طریق یک دگرگونی پیچیده ویژگی های یک حیوان بالغ را به دست می آورد. گذار از توسعه از طریق دگردیسی به توسعه مستقیم یکی از مهم ترین نتایج آخرین مراحل تکامل حیات روی زمین است.
علیرغم تقسیم بندی پیچیده افراد در درختان، بوته ها و علف های چند ساله، از نظر سطح سازماندهی انتوژنز آنها نسبت به گیاهان گلدار یک ساله، دو ساله و زودگذر پایین تر هستند. در دومی، انتوژنز با هماهنگی دقیق فعالیت زندگی تعداد معینی از اندام ها ادامه می یابد. فرآیندهای تمایز و مورفوژنز در انتوژنز آنها ماهیت "انفجاری" دارند.
در گیاهان، به دلیل رشد ضعیف سیستم تنظیمی، انتوژن با پایداری بیشتر مشخص می شود (به زیر مراجعه کنید). آنتوژنز در گیاهان به طور کلی بیشتر به شرایط محیطی بستگی دارد تا در حیوانات.
ویژگی های مشترک آنتوژنز در ارگانیسم های مختلف برنامه ریزی آن، جهت تمایز آن، توالی تغییرات در برنامه های توسعه تحت تأثیر عوامل محیطی (عوامل اپی ژنتیک) است.
تنوع آنتوژنز در گروه های مختلف ارگانیسم ها (حتی در میان نمایندگان یک گونه) نشان دهنده نقش ویژه عوامل محیطی در تثبیت تمایز و چرخه های زندگی است. اگرچه انتخاب در امتداد یک رویه زایی کل نگر رخ می دهد، مراحل فردی آن به عنوان پیش نیازهای لازم برای اجرای کل برنامه و جریان اطلاعات بین نسل ها عمل می کند.
در نمایندگان پادشاهی های مختلف، انواع، طبقات، آنتوژنز نیز در مقیاس تمایز متفاوت است. در موجودات تک سلولی از نظر پیچیدگی فرآیندهای تمایز اولیه است. در گیاهان، فرآیندهای تمایز گسترش یافته و به دوره رشد جنینی محدود نمی شود (تشکیل اندام های متامریک در گیاهان در طول انتوژن رخ می دهد). در حیوانات، فرآیندهای تمایز و تشکیل اندام عمدتاً به دوره جنینی محدود می شود. فرآیندهای هیستو و مورفوژنز در گیاهان پیچیده تر است و اندام ها و ساختارهای کمتری را نسبت به حیوانات درگیر می کند.
مدت زمان انتوژنز.در نمایندگان انواع مختلف، طبقات، سفارشات، مدت زمان انتوژنز یک ویژگی مهم گونه است. محدودیت امید به زندگی با شروع مرگ طبیعی، حتی در صورت وجود شرایط مساعد خارجی، یک نتیجه مهم از تکامل است که امکان تغییر نسل ها را فراهم می کند. در موجودات تک سلولی، انتوژنز با تشکیل سلولهای دختر پایان مییابد؛ مرگ از نظر مورفولوژیکی ثابت نیست (و آنها به معنای خاصی جاودانه هستند). در قارچ ها و گیاهان، پیری اندام های مختلف به طور نابرابر اتفاق می افتد. در قارچ، خود "میسلیوم" برای مدت طولانی در بستر زندگی می کند (در قارچ عسل علفزار (Marasmius oreades) - تا 500 سال!). از طرفی در بین قارچ ها موجودات زودگذری وجود دارند که هفته ها و ماه ها زندگی می کنند (Clavaria gyromitra). روی میز جدول 14.1 برخی از داده ها را در مورد امید به زندگی تعدادی از گیاهان نشان می دهد. گیاهان نیز مانند حیوانات از نظر طول عمر بسیار متنوع هستند.
| جدول 14.1. مدت زمان انتوژنز برخی از گونه ها | |
| انواع | مدت زمان آنتوژنز |
| 1. پادشاهی پیش از هسته ای | |
| سیانی | چند ساعت |
| II. پادشاهی قارچ | |
| پنی سیلیوم نوتاتوم | چند هفته |
| پولیپور (Fomes fomentarius) | تا 25 سال |
| قارچ سفید (Botulus botulus) | چند سال |
| III. پادشاهی گیاهی | |
| برش (Arabidopsis thaliana) | 60-70 روز |
| گندم (Triticum bulgare) | حدود 1 سال |
| انگور (Vitis vinifera) | 80-100 سال |
| درخت سیب (Malus domestica) | 200 سال |
| گردو (Juglans regia) | 300-400 سال |
| لیندن (Tilia grandifolia) | 1000 سال |
| بلوط (Quercus robur) | 1200 سال |
| سرو (Cupressus fastigiata) | 3000 سال |
| درخت ماموت (Sequoia gigantea) | 5000 سال |
| IV. پادشاهی حیوانات | |
| کرم نواری پهن (Diphyllobothrium latum) | تا 29 سال سن |
| مورچه (Formica fusca) | تا 7 سال |
| زنبور عسل (Apis mellifera) | تا 5 سال |
| خارپشت دریایی (Ehinus esculentus) | تا 8 سال |
| Com (Silurus glanis) | تا 60 سال سن |
| گوبی (Aphya pellucida) | 1 سال |
| وزغ معمولی (Bufo bufo) | تا 36 سال سن |
| لاک پشت (Testudo sumelri) | تا 150 سال |
| جغد عقاب معمولی (Bubo bubo) | تا 68 سال سن |
| کبوتر سنگی (Columba livid) | تا 30 سال سن |
| فیل آفریقایی (Elephas maximus) | تا 60 سال سن |
| گیبون (Hylobates lar) | تا 32 سال سن |
آنتوژنز- رشد فردی یک فرد، کلیت دگرگونی های به هم پیوسته آن که به طور طبیعی در طول چرخه زندگی از لحظه شکل گیری زیگوت تا مرگ رخ می دهد.
در حیوانات چند سلولی که از طریق جنسی تولید مثل می کنند، انتوژنز به دو دسته تقسیم می شود جنینی(از تشکیل زیگوت تا تولد یا بیرون آمدن از غشای تخمک) و پس از جنینیدوره های (از خروج از غشای تخمک یا تولد تا مرگ ارگانیسم). یک زیگوت در نتیجه ادغام سلول های تولید مثل نر و ماده - گامت ها تشکیل می شود. گامت ها بسته به ارگانیسم نر یا ماده در غدد جنسی تشکیل می شوند. فرآیند توسعه گامت نامیده می شود گامتوژنز. فرآیند تشکیل اسپرم نامیده می شود اسپرم زایی، و تشکیل تخم است اووژنز.
اسپرماتوژنز
1 - مرحله تولید مثل؛ 2 - مرحله رشد; 3 - مرحله بلوغ; 4 - مرحله تشکیل.
اسپرماتوژنز در بیضه ها اتفاق می افتد و به چهار مرحله تقسیم می شود: 1) تولید مثل، 2) رشد، 3) بلوغ، 4) تشکیل. در طول مرحله تولید مثل، اسپرماتوگونی دیپلوئید به طور مکرر توسط میتوز تقسیم می شود. برخی از اسپرماتوگونی های حاصل ممکن است تحت تقسیمات میتوزی مکرر قرار گیرند و در نتیجه همان سلول های اسپرماتوگونی تشکیل شوند. قسمت دیگر تقسیم را متوقف می کند و اندازه آن افزایش می یابد و وارد فاز بعدی اسپرماتوژنز می شود - مرحله رشد.
فاز رشد مربوط به فاز 1 میوز است، یعنی. در طی این فرآیند، سلول ها برای میوز آماده می شوند. رویداد اصلی مرحله رشد همانندسازی DNA است. در طول مرحله بلوغ، سلول ها توسط میوز تقسیم می شوند. در طول اولین تقسیم میوز آنها نامیده می شوند اسپرماتوسیت های درجه یک، در طول دوم - اسپرماتوسیت های درجه 2. از یک اسپرماتوسیت مرتبه اول، چهار اسپرماتید هاپلوئید به وجود می آید. مرحله تشکیل با این واقعیت مشخص می شود که اسپرماتیدهای کروی در ابتدا تحت یک سری دگرگونی های پیچیده قرار می گیرند که در نتیجه آن اسپرم ها تشکیل می شوند. تمام عناصر هسته و سیتوپلاسم در آن شرکت می کنند.
در انسان، اسپرم زایی در دوران بلوغ آغاز می شود. دوره تشکیل اسپرم سه ماه است، یعنی. اسپرم هر سه ماه یکبار تجدید می شود. اسپرماتوژنز به طور مداوم و همزمان در میلیون ها سلول اتفاق می افتد.

1 - "سر"؛ 2 - "گردن"؛ 3 - قسمت میانی؛ 4 - تاژک؛ 5 - آکروزوم; 6 - هسته؛ 7 - سانتریول؛ 8- میتوکندری.
اسپرم پستانداران به شکل یک نخ بلند است. طول اسپرم انسان 50-60 میکرون است. ساختار اسپرم را می توان به "سر"، "گردن"، یک بخش میانی و یک دم تقسیم کرد. سر شامل هسته و آکروزوم است. هسته شامل مجموعه ای از کروموزوم هاپلوئید است. آکروزوم یک اندامک غشایی حاوی آنزیم هایی است که برای حل کردن غشای تخم استفاده می شود. دو سانتریول در گردن و میتوکندری در بخش میانی وجود دارد. دم با یک، در برخی گونه ها - دو یا چند تاژک نشان داده می شود. تاژک اندامک حرکتی است و از نظر ساختار شبیه تاژک ها و مژک های تک یاخته است. برای حرکت تاژک ها از انرژی پیوندهای ماکرو ارژیک ATP استفاده می شود؛ سنتز ATP در میتوکندری اتفاق می افتد.
اسپرم در سال 1677 توسط A. Leeuwenhoek کشف شد.
اووژنز
در تخمدان ها انجام می شود و به سه مرحله تقسیم می شود: 1) تولید مثل، 2) رشد، 3) بلوغ.
در طول فاز تولید مثل، اوگونیاهای دیپلوئیدی به طور مکرر توسط میتوز تقسیم می شوند. فاز رشد مربوط به فاز 1 میوز است، یعنی. در طی آن، سلول ها برای میوز آماده می شوند: اندازه سلول ها به دلیل تجمع مواد مغذی به طور قابل توجهی افزایش می یابد. رویداد اصلی مرحله رشد همانندسازی DNA است. در طول مرحله بلوغ، سلول ها با میوز تقسیم می شوند. در طول اولین تقسیم میوز آنها نامیده می شوند تخمک های مرتبه 1. در نتیجه اولین تقسیم میوز، دو سلول دختر به وجود می آیند: یک سلول کوچک، به نام اولین بدن قطبی، و بزرگتر - تخمک مرتبه 2. در طول تقسیم میوز دوم، تخمک مرتبه 2 تقسیم می شود تا تخم و جسم قطبی دوم را تشکیل دهد و جسم قطبی اول تقسیم می شود و اجسام قطبی سوم و چهارم را تشکیل می دهد. بنابراین، در نتیجه میوز، یک تخمک و سه جسم قطبی از یک تخمک درجه یک تشکیل می شود.

1-مرحله تولید مثل؛ 2 - مرحله رشد; 3 - مرحله بلوغ.
بر خلاف تشکیل اسپرم که تنها پس از رسیدن به بلوغ اتفاق می افتد، فرآیند تشکیل تخمک در انسان از دوره جنینی آغاز می شود و به طور متناوب ادامه می یابد. در جنین مراحل تولید مثل و رشد به طور کامل محقق می شود و مرحله بلوغ آغاز می شود. تا زمانی که یک دختر متولد می شود، تخمدان های او حاوی صدها هزار تخمک مرتبه اول هستند که در مرحله دیپلوتن پروفاز 1 میوز متوقف شده و "یخ زده" شده اند. بلوک اول اووژنز.
در دوران بلوغ، میوز از سر گرفته می شود: تقریباً هر ماه، تحت تأثیر هورمون های جنسی، یکی از تخمک ها (به ندرت دو تخمک) به متافاز 2 میوز می رسد - بلوک دوم اووژنز. میوز تنها در صورتی می تواند تکمیل شود که لقاح رخ دهد. اگر لقاح اتفاق نیفتد، تخمک مرتبه دوم می میرد و از بدن دفع می شود.
ساختار تخم مرغ
شکل تخم ها معمولا گرد است. اندازه تخم مرغ ها بسیار متفاوت است - از چند ده میکرومتر تا چند سانتی متر (تخم مرغ انسان حدود 120 میکرون است). ویژگیهای ساختاری تخمها عبارتند از: وجود غشاهای واقع در بالای غشای پلاسمایی و وجود مقدار کم و بیش زیادی از مواد مغذی ذخیره در سیتوپلاسم.

1 - پیش هسته در متا فاز 2. 2 - پوسته براق؛ 3-پوسته تابشی 4- جسم قطبی اول.
در بیشتر حیوانات، تخمها دارای غشای اضافی هستند که در بالای غشای سیتوپلاسمی قرار دارند. بسته به مبدا، آنها متمایز می شوند: پوسته های اولیه، ثانویه و سوم. پوسته های اولیهاز مواد ترشح شده توسط تخمک تشکیل می شوند. لایه ای در تماس با غشای سیتوپلاسمی تخمک تشکیل می شود. این یک عملکرد محافظتی انجام می دهد، ویژگی گونه های نفوذ اسپرم را تضمین می کند، یعنی. از نفوذ اسپرم گونه های دیگر به تخمک جلوگیری می کند. در پستانداران به این غشاء پلوسیدا می گویند. پوسته های ثانویهاز ترشح سلول های فولیکولی تخمدان تشکیل می شوند و در همه تخمک ها وجود ندارند. پوسته ثانویه تخم حشرات حاوی یک کانال - میکروپیل است که از طریق آن اسپرم به داخل تخمک نفوذ می کند. پوسته های درجه سومبه دلیل فعالیت غدد خاص مجاری تخمک تشکیل می شوند. به عنوان مثال، از ترشحات غدد خاص، غشاهای پروتئین، پوسته زیر پوسته، پوسته و غشای فوق پوسته در پرندگان و خزندگان تشکیل می شود.
غشاهای ثانویه و سوم در تخم های حیوانی تشکیل می شوند که جنین های آن در محیط بیرونی رشد می کنند. از آنجایی که پستانداران تحت رشد داخل رحمی قرار میگیرند، تخمهای آنها فقط یک غشای اولیه دارند که در بالای آن تاج رادیاتا قرار دارد، لایهای از سلولهای فولیکولی که مواد مغذی را به تخم میرسانند.
در تخم مرغ، مواد مغذی انباشته می شود که به آنها گفته می شود زرده. حاوی چربی ها، کربوهیدرات ها، RNA، مواد معدنی، پروتئین ها است که بخش عمده آن لیپوپروتئین و گلیکوپروتئین است. زرده در سیتوپلاسم موجود است دانه های زرده. مقدار مواد مغذی انباشته شده در تخمک بستگی به شرایطی دارد که جنین در آن رشد می کند. اگر رشد تخم در خارج از بدن مادر اتفاق بیفتد و منجر به تشکیل حیوانات بزرگ شود، زرده می تواند بیش از 95٪ از حجم تخم مرغ را تشکیل دهد. تخمهای پستاندارانی که در داخل بدن مادر رشد میکنند حاوی مقدار کمی زرده هستند - کمتر از 5٪، زیرا جنینها مواد مغذی لازم برای رشد را از مادر دریافت میکنند.

1 - السیتال; 2 - ایزوله-سیتال; 3 - تللسیتال متوسط; 4 - تلولسیتال شدید.
بسته به مقدار زرده موجود، موارد زیر مشخص می شوند: انواع تخم مرغ: alecital(حاوی زرده نباشد یا دارای مقدار کمی زرده باشد - پستانداران، کرم های مسطح). ایزوولسیتال(با زرده به طور مساوی توزیع شده - نیزه، خارپشت دریایی)؛ تللسیتالی متوسط(با زرده توزیع نابرابر - ماهی، دوزیستان)؛ به شدت تلوسیتال(زرده بیشترین قسمت را اشغال می کند و فقط قسمت کوچکی از سیتوپلاسم در قطب حیوان از آن آزاد است - پرندگان).
به دلیل تجمع مواد مغذی، تخم ها قطبیت پیدا می کنند. قطب مخالف نامیده می شود رویشیو حیوانی. پلاریزاسیون در این واقعیت آشکار می شود که محل هسته در سلول تغییر می کند (به سمت قطب حیوان جابجا می شود) و همچنین در توزیع آخال های سیتوپلاسمی (در بسیاری از تخم مرغ ها مقدار زرده از حیوان افزایش می یابد. به قطب نباتی).
تخم انسان در سال 1827 توسط K.M. بائر.
لقاح
فرآیند ادغام سلولهای تولیدمثلی نر و ماده، که منجر به تشکیل زیگوت میشود که ارگانیسم جدیدی را ایجاد میکند، نامیده میشود. لقاح. خود فرآیند لقاح از لحظه تماس اسپرم و تخمک آغاز می شود. در لحظه چنین تماسی، غشای پلاسمایی رشد آکروزوم و قسمت مجاور غشای وزیکول آکروزوم حل می شود، آنزیم هیالورونیداز و سایر مواد فعال بیولوژیکی موجود در آکروزوم آزاد می شود و بخشی از غشای تخم مرغ را حل می کند. . اغلب، اسپرم به طور کامل به داخل تخمک جمع می شود، گاهی اوقات تاژک بیرون می ماند و دور ریخته می شود. از لحظه ای که اسپرم به تخمک نفوذ می کند، گامت ها از بین می روند، زیرا آنها یک سلول واحد - زیگوت را تشکیل می دهند. هسته اسپرم متورم می شود، کروماتین آن شل می شود، غشای هسته حل می شود و تبدیل به پیش هسته نر. این امر همزمان با تکمیل دومین تقسیم میوزی هسته تخمک اتفاق میافتد که به دلیل لقاح دوباره شروع شد. به تدریج هسته تخمک تبدیل می شود پیش هسته ماده. پیش هسته ها به سمت مرکز تخمک حرکت می کنند، همانندسازی DNA اتفاق می افتد و پس از همجوشی آنها، مجموعه کروموزوم ها و DNA زیگوت تبدیل به "2" می شود. n 4ج" اتحاد پیش هسته ها نشان دهنده لقاح واقعی. بنابراین، لقاح با تشکیل یک زیگوت با یک هسته دیپلوئید به پایان می رسد.
لقاح یک فرآیند برگشت ناپذیر است، به این معنی که یک تخمک پس از بارور شدن نمی تواند دوباره بارور شود.
بسته به تعداد افرادی که در تولید مثل جنسی شرکت می کنند، عبارتند از: لقاح متقابل- لقاح، که در آن گامت های تشکیل شده توسط ارگانیسم های مختلف شرکت می کنند. خود باروری- لقاح، که در آن گامت های تشکیل شده توسط همان ارگانیسم (کرم های نواری) ادغام می شوند.
دوره جنینی
تقسیم کردن
تقسیم کردنمجموعه ای از تقسیمات میتوزی متوالی زیگوت است که در نتیجه حجم عظیمی از سیتوپلاسم تخمک به سلول های کوچکتر متعدد حاوی هسته تقسیم می شود. در نتیجه تکه تکه شدن، سلول هایی تشکیل می شوند که بلاستومر نامیده می شوند. چیزی که برش را از تقسیم معمولی متمایز می کند این است که بلاستومرهای تازه تشکیل شده از نظر اندازه افزایش نمی یابند. این به دلیل از دست دادن دوره پیش سنتز اینترفاز ممکن می شود. در این مورد، دوره مصنوعی اینترفاز در تلوفاز میتوز قبلی شروع می شود. بنابراین، تعداد بلاستومرها به تدریج افزایش می یابد، اما حجم کل آنها تقریباً بدون تغییر باقی می ماند. سیتوپلاسم سلول ها در حین تکه تکه شدن با وقوع هجوم غشای سلولی تقسیم می شود. شیارهای خرد کردن).
شکاف تخم دوزیستان (قورباغه): 1 - مرحله دو سلولی; 2 - مرحله چهار سلولی; 3 - مرحله هشت سلولی;
4- گذار از مرحله هشت سلولی به شانزده سلولی (سلولهای قطب حیوانی قبلاً تقسیم شده اند و سلولهای قطب رویشی.
تازه شروع به تکه تکه شدن) 5 - مرحله بعدی خرد کردن. 6 - بلاستولا; 7 - بلاستولا در مقطع.
اهمیت بیولوژیکی فرآیند خرد کردن: به لطف تکرار چرخه های تولید مثل، ژنوتیپ زیگوت چند برابر می شود. توده سلولی برای دگرگونی های بیشتر جمع می شود؛ جنین از تک سلولی به چند سلولی تبدیل می شود.
تقسیم بلاستومر رخ می دهد همزمانو غیر همزمان. در اکثر گونه ها از همان ابتدای رشد ناهمزمان است، در سایر گونه ها پس از اولین تقسیمات ناهمزمان می شود.
ماهیت تکه تکه شدن، اول از همه، با ساختار تخم مرغ، عمدتاً با مقدار زرده و ویژگی های توزیع آن در سیتوپلاسم تعیین می شود. در این راستا، با توجه به روش خرد کردن، دو نوع اصلی تخم مرغ را تشخیص می دهند: کاملا له شده و نیمه له شده. شکاف کامل زمانی نامیده می شود که سیتوپلاسم تخمک به طور کامل به بلاستومرها تقسیم شود. ممکن است لباس فرم- همه بلاستومرهای تشکیل شده اندازه و شکل یکسانی دارند (معمولی برای تخم های آلسیتال و ایزولسیتال) و ناهموار. ناجور- بلاستومرهایی با اندازه نابرابر تشکیل می شوند (معمولاً تخم مرغ های تلولسیتال با محتوای زرده متوسط). بلاستومرهای کوچک در قطب حیوان، بزرگ - در ناحیه قطب رویشی جنین بوجود می آیند.

تکمیل شده؛ ب - جزئی؛ ب - دیسکی.
خرد کردن جزئی- نوعی شکاف که در آن سیتوپلاسم تخمک به طور کامل به بلاستومرها تقسیم نمی شود. یکی از انواع شکاف جزئی دیسکوئیدی است که در آن تنها بخش بدون زرده سیتوپلاسم در قطب حیوانی که هسته در آن قرار دارد، دچار شکاف می شود. ناحیه ای از سیتوپلاسم که دچار تکه تکه شدن شده است نامیده می شود دیسک ژرمینال. این نوع خرد کردن برای تخم مرغ های تللسیتال قوی با مقدار زیادی زرده (خزندگان، پرندگان، ماهی) معمول است.
تکه تکه شدن در نمایندگان گروه های مختلف حیوانات ویژگی های خاص خود را دارد، اما با تشکیل ساختاری مشابه - بلاستولا - به پایان می رسد.
بلاستولا
بلاستولا- جنین تک لایه از لایه ای از سلول ها تشکیل شده است - بلاستودرم که حفره را محدود می کند - بلاستوکول. بلاستولا در مراحل اولیه شکاف به دلیل واگرایی بلاستومرها شروع به تشکیل می کند. حفره حاصل با مایع پر می شود. ساختار بلاستولا تا حد زیادی به نوع شکاف بستگی دارد.
کولوبلاستولا(بلاستولای معمولی) با تکه تکه شدن یکنواخت تشکیل می شود. به نظر می رسد یک وزیکول تک لایه با یک بلاستوکول بزرگ (لنسلت).
آمفیبلاستولااز خرد کردن تخمهای تلولسیتال تشکیل میشود. بلاستودرم از بلاستومرهایی با اندازه های مختلف ساخته شده است: میکرومرها در قطب حیوانی و ماکرومرها در قطب های رویشی. در این حالت بلاستوکول به سمت قطب حیوانی (دوزیستان) جابه جا می شود.

1 - کولوبلاستولا؛ 2 - آمفیبلاستولا; 3 - دیسکوبلاستولا; 4 - بلاستوسیست; 5 - جنین. 6 - تروفوبلاست.
دیسکوبلاستولااز خرد کردن دیسکی شکل تشکیل شده است. حفره بلاستولا شبیه شکاف باریکی است که در زیر دیسک ژرمینال (پرنده) قرار دارد.
بلاستوسیستیک وزیکول تک لایه پر از مایع است که در آن یک جنین (که جنین از آن رشد می کند) و یک تروفوبلاست وجود دارد که تغذیه جنین (پستانداران) را تامین می کند.

1 - اکتودرم؛ 2 - اندودرم؛ 3 - بلاستوپور; 4 - گاستروکول.
پس از تشکیل بلاستولا، مرحله بعدی جنین زایی آغاز می شود - گاسترولاسیون(تشکیل لایه های جوانه). در نتیجه گاسترولاسیون، یک جنین دو لایه و سپس یک جنین سه لایه (در اکثر حیوانات) تشکیل می شود - گاسترولا. در ابتدا، لایه های بیرونی (اکتودرم) و داخلی (اندودرم) تشکیل می شوند. بعداً، لایه سوم زاینده، مزودرم، بین اکتو و اندودرم تشکیل می شود.
لایه های جوانه- لایه های جدا از سلول ها که موقعیت خاصی را در جنین اشغال می کنند و اندام ها و سیستم های اندام مربوطه را ایجاد می کنند. لایه های جوانه نه تنها در نتیجه حرکت توده های سلولی، بلکه در نتیجه تمایز سلول های بلاستولای مشابه و نسبتا همگن ایجاد می شوند. در طی فرآیند گاسترولاسیون، لایه های جوانه موقعیتی را اشغال می کنند که مطابق با طرح ساختاری ارگانیسم بالغ است. تفکیک- روند ظهور و افزایش تفاوت های مورفولوژیکی و عملکردی بین سلول های فردی و بخش هایی از جنین. بسته به نوع بلاستولا و ویژگی های حرکت سلولی، روش های اصلی گاسترولاسیون زیر مشخص می شود: انواژشن، مهاجرت، لایه برداری، اپیبولی.

1 - انواژیناسیون; 2 - اپیبولیک؛ 3 - مهاجرت; 4 - لایه لایه شدن;
الف - اکتودرم؛ ب - اندودرم؛ ج - گاستروکول.
در انواژیناسیونیکی از بخش های بلاستودرم شروع به نفوذ به داخل بلاستوکول (در لنسلت) می کند. در این حالت، بلاستوکول تقریباً به طور کامل جابجا می شود. یک کیسه دو لایه تشکیل می شود که دیواره خارجی آن اکتودرم اولیه و دیواره داخلی آندودرم اولیه است که حفره روده اولیه را می پوشاند یا گاستروسل. سوراخی که از طریق آن حفره با محیط ارتباط برقرار می کند نامیده می شود بلاستوپور، یا دهان اولیه. در نمایندگان گروه های مختلف حیوانات، سرنوشت بلاستوپور متفاوت است. در پروتوستوم ها به دهانه دهان تبدیل می شود. در دوتروستوم ها، بلاستوپور بیش از حد رشد می کند و در جای آن اغلب یک دهانه مقعدی ظاهر می شود و دهانه دهان در قطب مخالف (انتهای قدامی بدن) می شکند.
مهاجرت- "اخراج" بخشی از سلول های بلاستودرم به داخل حفره بلاستوکول (مهره داران بالاتر). از این سلول ها اندودرم تشکیل می شود.
لایه لایه شدگیدر حیواناتی که دارای بلاستولای بدون بلاستوکول (پرندگان) هستند رخ می دهد. با این روش گاسترولاسیون، حرکات سلولی حداقل یا کاملاً وجود ندارد، زیرا لایه بندی رخ می دهد - سلول های بیرونی بلاستولا به اکتودرم تبدیل می شوند و سلول های داخلی آندودرم را تشکیل می دهند.
اپیبولیزمانی اتفاق میافتد که بلاستومرهای کوچکتر قطب حیوانی سریعتر تکه تکه میشوند و بلاستومرهای بزرگتر قطب گیاهی بیش از حد رشد میکنند و اکتودرم (دوزیستان) را تشکیل میدهند. سلول های قطب رویشی باعث ایجاد لایه ژرم داخلی یعنی اندودرم می شود.
روش های توصیف شده گاسترولاسیون به ندرت به صورت خالص یافت می شوند و معمولاً ترکیبات آنها مشاهده می شود (درهم رفتگی با اپیبولی در دوزیستان یا لایه برداری با مهاجرت در خارپوستان).
اغلب، مواد سلولی مزودرم بخشی از اندودرم است. به شکل برآمدگی های جیبی شکل به داخل بلاستوکول نفوذ می کند و سپس جدا می شود. هنگامی که مزودرم تشکیل می شود، یک حفره بدن ثانویه یا کوئلوم تشکیل می شود.
فرآیند تشکیل اندام در رشد جنینی نامیده می شود اندام زایی. اندام زایی را می توان به دو مرحله تقسیم کرد: عصبی سازی- تشکیل مجموعه ای از اندام های محوری (لوله عصبی، نوتوکورد، لوله روده و مزودرم سومیت) که تقریباً کل جنین را درگیر می کند، و ساخت سایر اندام ها، به دست آوردن توسط قسمت های مختلف بدن از شکل معمولی و ویژگی های سازمان داخلی، ایجاد نسبت های خاص (فرایندهای محدود فضایی).
توسط نظریه لایه جوانه کارل بائر، ظهور اندام ها به دلیل تبدیل یک یا یک لایه جوانه دیگر - اکتو-، مزو- یا اندودرم است. برخی از اندام ها ممکن است منشا مختلط داشته باشند، یعنی با مشارکت چندین لایه جوانه به طور همزمان تشکیل می شوند. برای مثال ماهیچه های دستگاه گوارش مشتق مزودرم و پوشش داخلی آن مشتق از آندودرم است. با این حال، تا حدودی ساده تر، منشاء اندام های اصلی و سیستم های آنها هنوز می تواند با لایه های جوانه خاصی مرتبط باشد. جنین در مرحله نورولاسیون نامیده می شود نورولا. مواد مورد استفاده برای ساختن سیستم عصبی در مهره داران - نورواکتودرم، بخشی از قسمت پشتی اکتودرم است. در بالای ریشه نوتوکورد قرار دارد.

1 - اکتودرم؛ 2 - وتر; 3 - حفره ثانویه بدن; 4 - مزودرم; 5 - اندو درم؛ 6 - حفره روده؛ 7 - لوله عصبی.
ابتدا در ناحیه نورواکتودرم، صاف شدن لایه سلولی رخ می دهد که منجر به تشکیل صفحه عصبی می شود. سپس لبه های صفحه عصبی ضخیم شده و بالا می روند و چین های عصبی را تشکیل می دهند. در مرکز صفحه، به دلیل حرکت سلول ها در امتداد خط وسط، یک شیار عصبی ظاهر می شود که جنین را به دو نیمه راست و چپ آینده تقسیم می کند. صفحه عصبی در امتداد خط وسط شروع به تا شدن می کند. لبه های آن لمس می شود و سپس بسته می شود. در نتیجه این فرآیندها، یک لوله عصبی با یک حفره ظاهر می شود - neurocoelom.
بسته شدن برجستگی ها ابتدا در وسط و سپس در قسمت خلفی شیار عصبی رخ می دهد. در نهایت، این اتفاق در قسمت سر، که پهن تر از بقیه است، رخ می دهد. بخش منبسط شده قدامی بیشتر مغز را تشکیل می دهد، بقیه لوله عصبی نخاع را تشکیل می دهد. در نتیجه صفحه عصبی به یک لوله عصبی در زیر اکتودرم تبدیل می شود.
در طول نورولاسیون، برخی از سلول های صفحه عصبی بخشی از لوله عصبی نیستند. آنها صفحه گانگلیون یا تاج عصبی را تشکیل می دهند، مجموعه ای از سلول ها در امتداد لوله عصبی. بعداً این سلولها در سراسر جنین مهاجرت میکنند و سلولهای عقدههای عصبی، مدولای آدرنال، سلولهای رنگدانه و غیره را تشکیل میدهند.
از مواد اکتودرم، علاوه بر لوله عصبی، اپیدرم و مشتقات آن (پر، مو، ناخن، پنجه، غدد پوست و غیره)، اجزای اندام های بینایی، شنوایی، بویایی، اپیتلیوم دهان و... مینای دندان رشد می کند.
اندام های مزودرمی و اندودرمی نه پس از تشکیل لوله عصبی، بلکه همزمان با آن تشکیل می شوند. در امتداد دیواره های جانبی روده اولیه، جیب ها یا چین ها با بیرون زدگی اندودرم تشکیل می شوند. ناحیه آندودرم که بین این چین ها قرار دارد ضخیم می شود، خم می شود، چین می خورد و از توده اصلی آندودرم جدا می شود. اینگونه ظاهر می شود وتر. برآمدگی های پاکتی مانند آندودرم از روده اولیه جدا شده و به یک سری کیسه های بسته که به صورت قطعه ای قرار گرفته اند تبدیل می شوند که کیسه های سلومیک نیز نامیده می شوند. دیواره های آنها توسط مزودرم تشکیل شده است و حفره داخل یک حفره بدن ثانویه است (یا به طور کلی).
انواع بافت همبند، درم، اسکلت، ماهیچه های مخطط و صاف، سیستم گردش خون و لنفاوی و دستگاه تناسلی از مزودرم ایجاد می شوند.
از اندودرم، اپیتلیوم روده ها و معده، سلول های کبد، سلول های ترشح کننده لوزالمعده، روده و غدد معده رشد می کنند. بخش قدامی روده جنینی اپیتلیوم ریه ها و راه های هوایی را تشکیل می دهد و بخش هایی از لوب های قدامی و میانی غده هیپوفیز، تیروئید و غدد پاراتیروئید را ترشح می کند.

1 - ریشه کوردو-مزودرم; 2 - حفره بلاستولا; 3 - لوله عصبی القا شده. 4 - وتر القایی; 5 - لوله عصبی اولیه; 6 - وتر اولیه؛ 7- تشکیل جنین ثانویه متصل به جنین میزبان.
- این یک تعامل بین بخش هایی از جنین است که در طی آن یک قسمت از آن - القاء کننده - تماس با قسمت دیگر - سیستم واکنش - جهت رشد دومی را تعیین می کند.
پدیده القاء توسط H. Spemann در سال 1901 هنگام مطالعه تشکیل عدسی چشم از اپیتلیوم اکتودرمی در جنین های دوزیستان کشف شد. در سال 1924، نتایج آزمایش های H. Spemann و G. Mangold منتشر شد که اثبات کلاسیک وجود القای جنینی در نظر گرفته شد. در مرحله گاسترولای اولیه، پایه اکتودرم، که در شرایط عادی باید به ساختارهای سیستم عصبی تبدیل می شد، از جنین نیوت تاج دار (غیر رنگدانه) زیر اکتودرم سمت شکم پیوند زده شد. ایجاد اپیدرم پوست، جنین نیوت معمولی (رنگدانه دار). در نتیجه، لوله عصبی و سایر اجزای کمپلکس اندام محوری ابتدا در سمت شکمی جنین گیرنده ظاهر شد و سپس یک جنین اضافی تشکیل شد. علاوه بر این، مشاهدات نشان داده است که بافتهای جنین اضافی تقریباً منحصراً از مواد سلولی گیرنده تشکیل میشوند.
اگر ریشه نوتوکورد در مرحله اولیه گاسترول به طور کامل برداشته شود، لوله عصبی رشد نمی کند. اکتودرم در سمت پشتی جنین، که لوله عصبی به طور معمول از آن تشکیل می شود، اپیتلیوم پوست را تشکیل می دهد. پس از مطالعه بیشتر در مورد رشد جنین ها، مشخص شد که رودیمنت کوردومزودرم، که یک القاء کننده لوله عصبی است، برای تمایز نیاز به تأثیر القایی از پایه سیستم عصبی دارد.
دوره رشد پس از جنین
دوره رشد پس از جنین از لحظه تولد یا رها شدن ارگانیسم از غشای تخم شروع می شود و تا زمان مرگ ادامه می یابد. رشد پس از جنین شامل: رشد ارگانیسم. ایجاد تناسب نهایی بدن؛ انتقال سیستم های اندام به حالت یک ارگانیسم بالغ (به ویژه بلوغ). تمیز دادن دو نوع اصلی رشد پس از جنین: 1) مستقیم، 2) با دگرگونی.
در توسعه مستقیمیک فرد از بدن مادر یا پوسته تخم بیرون می آید و تنها در اندازه کوچکتر (پرندگان، پستانداران) با ارگانیسم بالغ متفاوت است.

وجود دارد: غیر لارونوع (تخمزا)، که در آن جنین در داخل تخمک (ماهی، پرنده) رشد میکند. داخل رحمینوعی که در آن جنین در داخل بدن مادر رشد می کند و از طریق جفت (پستانداران جفتی) به آن متصل می شود.
در طی رشد با دگرگونی (دگردیسی)، یک لارو از تخم بیرون میآید که از نظر ساختار سادهتر از یک حیوان بالغ است (گاهی اوقات بسیار متفاوت از آن). به عنوان یک قاعده، اندام های لارو خاصی دارد و اغلب سبک زندگی متفاوتی نسبت به یک حیوان بالغ (حشرات، برخی از عنکبوتیان، دوزیستان) دارد.
به عنوان مثال، در دوزیستان بدون دم، یک لارو، یک قورباغه، از پوسته تخم بیرون می آید. شکل بدنی صاف، باله دمی، شکاف ها و آبشش های آبشش، اندام های خط جانبی، قلب دو حفره ای و یک گردش خون دارد. با گذشت زمان، تحت تأثیر هورمون تیروئید، قورباغه دچار دگردیسی می شود. دم او حل می شود، اندام ظاهر می شود، خط جانبی ناپدید می شود، ریه ها و دایره دوم گردش خون ایجاد می شود، یعنی. به تدریج ویژگی های مشخصه دوزیستان را به دست می آورد.
پارتنوژنز
پارتنوژنزبه نام رشد یک ارگانیسم از یک تخمک بارور نشده. در تعدادی از گونه های گیاهی، بی مهرگان و مهره داران، به جز پستانداران، که در آنها جنین های پارتنوژنتیک در مراحل اولیه جنین زایی می میرند، یافت می شود. پارتنوژنز می تواند مصنوعی یا طبیعی باشد.
پارتنوژنز مصنوعیتوسط فرد با فعال کردن تخم مرغ با قرار دادن آن در معرض مواد مختلف، تحریک مکانیکی، افزایش دما و غیره ایجاد می شود.
در پارتنوژنز طبیعیتخمک شروع به تکه تکه شدن و تبدیل شدن به جنین بدون مشارکت اسپرم می کند، فقط تحت تأثیر علل داخلی یا خارجی. تمیز دادن جسمیو مولدپارتنوژنز در طول پارتنوژنز زایشی، یا هاپلوئید، جنین از یک تخم هاپلوئید (زنبورهای بدون سرنشین) شروع به رشد می کند. در پارتنوژنز سوماتیک یا دیپلوئید، جنین از یک سلول دیپلوئید شروع به رشد می کند: 1) یا از یک تخمک دیپلوئید (میوز رخ نمی دهد)، 2) یا از سلولی که در نتیجه ادغام دو هسته هاپلوئید (میوز) ایجاد شده است. رخ می دهد) (شته، دافنی، قاصدک).
اگر رشد تخمک بدون مشارکت هسته اسپرم (برخی ماهی ها، کرم های گرد) اتفاق بیفتد، این نوع پارتنوژنز نامیده می شود. ژنوژنز. با این حال، این اسپرم است که شروع تکه تکه شدن تخمک را تحریک می کند، اگرچه آن را بارور نمی کند.
اگر رشد تخمک فقط به دلیل ماده ژنتیکی اسپرم و سیتوپلاسم تخمک اتفاق بیفتد، در این مورد آنها صحبت می کنند. آندروژنز. این نوع رشد می تواند اتفاق بیفتد اگر هسته تخمک حتی قبل از لقاح بمیرد و نه یک، بلکه چند اسپرم وارد تخمک شوند (کرم ابریشم).
رفتن به سخنرانی شماره 15"تکثیر جنسی در گلدان"
رفتن به سخنرانی شماره 17"مفاهیم اساسی ژنتیک. قوانین مندل"
دوره ای شدن انتوژن موجودات چند سلولی
مرحله ژرمینال (جنین) و دوره های آن در حیوانات.
4. مرحله جنینیزمانی است که یک ارگانیسم جدید در داخل بدن مادر یا داخل تخمک رشد می کند. جنین زایی با تولد (جوانه زایی، جوانه زدن) به پایان می رسد. دوره جنینی پس از لقاح یا فعال شدن تخمک در طی پارتنوژنز آغاز می شود و در داخل بدن مادر، تخمک و دانه رخ می دهد. رشد جنینی با تولد (پستانداران)، بیرون آمدن از پوسته تخم مرغ (پرندگان، خزندگان) و جوانه زنی (گیاهان بذر) به پایان می رسد. مراحل اصلی دوره جنینی برش، گاسترولاسیون، هیستوژنز و اندامزایی است.
تقسیم کردن- مجموعه ای از تقسیمات میتوزی متوالی زیگوت که با تشکیل یک مرحله تک لایه - بلاستولا به پایان می رسد. تعداد سلول ها در نتیجه میتوز افزایش می یابد، اما اینترفاز بسیار کوتاه است و بلاستومرها رشد نمی کنند. خصوصیات له کردن در گروه های مختلف موجودات به ماهیت محل و مقدار زرده بستگی دارد که از این نظر دو نوع خردایش از هم متمایز می شود.
گاسترولاسیون - این فرآیند تشکیل یک جنین دو لایه - گاسترولا است. رشد سلولی در حین گاسترولاسیون اتفاق نمی افتد. در این مرحله، دو یا سه لایه از بدن جنین تشکیل می شود - لایه های جوانه. در فرآیند گاسترولاسیون، تمایز بین دو مرحله بسیار مهم است: الف) تشکیل اکتو و اندودرم (گاسترولای اولیه تشکیل می شود - جنین دو لایه) ب) تشکیل مزودرم (گاسترولای دیررس تشکیل می شود. - جنین سه لایه). در مرحله گاسترولاسیون، جنین زایی حیوانات دولایه (اسفنج ها، کولنترات ها) تکمیل می شود، مزودرم در رشد جنینی حیوانات سه لایه قرار می گیرد (شروع با کرم های مسطح).
در موجودات مختلف، گاسترولا به روش های مختلفی تشکیل می شود. انواع زیر از تشکیل گاسترول متمایز می شود: انواژیناسیون (انواژیناسیون)، لایه برداری (طبقه بندی)، اپیبولی (فولینگ)، مهاجرت (خزنده).
هیستوژنز و اندام زایی - تشکیل بافت ها و اندام ها. این فرآیندها به دلیل تمایز (ظهور تفاوت در ساختار و عملکرد سلول ها، بافت ها، اندام ها) انجام می شود. سلول های اولیه بافت های آموزشی در هیستوژنز گیاهان و سلول های بنیادی، نیمه ساقه ای و بالغ در هیستوژنز حیوانات شرکت می کنند. فعل و انفعالات بین سلولی و تأثیر مواد فعال بیولوژیکی نقش زیادی در اندام زایی دارند. مراحل هیستوژنز و اندام زایی (با استفاده از مثال لنسلت) عصبی شدن است - تشکیل مجموعه محوری اندام ها (لوله عصبی، نوتوکورد)، تشکیل سایر اندام ها - اندام ها ویژگی های ساختاری مشخصه بزرگسالان را به دست می آورند. اندام زایی عمدتاً در پایان دوره رشد جنینی کامل می شود، اما تمایز و عارضه اندام ها در پس از جنین زایی ادامه دارد.
دورهبندی انتوژنز موجودات چند سلولی - مفهوم و انواع. طبقه بندی و ویژگی های رده "پریودسازی انتوژنز ارگانیسم های چند سلولی" 2017، 2018.

")







- روند توسعه کشاورزی
- یاکولف الکساندر سرگیویچ ارزیابی خلاقیت در زمینه هوانوردی و با یاکولف
- شاهزاده ایزیاسلاو اول ایزیاسلاو در کدام شهر حکومت می کرد؟
- آب سیب در خانه - یک دستور العمل ساده بدون آبمیوه گیری
- چگونه تخم مرغ را در آرام پز، بخارپز، در آب و به عنوان املت بپزیم؟
- خیارهای زودپز خوشمزه در آب نمک کم نمک
- نحوه ترشی خیار کم نمک به روش های مختلف
- نحوه خاموش کردن سودا با سرکه برای پخت، چه نوع سرکه ای مورد نیاز است، چه چیزی می تواند جایگزین آن شود
- آب مرغ از مرغ خانگی
- چگونه یک فرد ناخواسته را از خانه دور کنیم؟
- Okroshka - تهیه غذا و ظروف
- خواص فیزیکی اکسیدها
- برگه تقلب: رشد فردی ارگانیسم ها (آنتوژنز)
- نکراسوف نیکولای الکسیویچ نقشه ای بسازید نیکولای الکسیویچ نکراسوف
- ساده ترین معادلات مثلثاتی روش های حل معادلات مثلثاتی
- حل معادلات مثلثاتی معرفی زاویه کمکی
- رابطه ژنتیکی بین طبقات مواد واکنش بین نمایندگان دو سری ژنتیکی
- دانشگاه فنی ملی پولتاوا به نام یو
- اطلاعات جالب در مورد ماه گرفتگی در چه فاز ماه می توان خورشید را رصد کرد؟
- آیا قبل از کلمه "چگونه" کاما وجود دارد یا خیر؟